Mga organo ng halaman: dahon. Mga kumplikadong inflorescences - sa isang karaniwang axis ay mga simpleng inflorescences. pataas - mga shoot na unang tumubo sa pahalang at pagkatapos ay patayong direksyon
Ang mga halaman ay tinatawag na mga photosynthetic na buhay na organismo na may kaugnayan sa mga eukaryotes. Mayroon silang cellular cellulose membrane, isang reserbang nutrient sa anyo ng starch, ay hindi aktibo o hindi kumikibo at lumalaki sa buong buhay nila. Ang chlorophyll pigment na taglay nito ay nagbibigay sa mga halaman ng kanilang berdeng kulay. Sa liwanag, mula sa carbon dioxide at tubig, lumilikha sila ng mga organikong sangkap at naglalabas ng oxygen, sa gayon ay nagbibigay ng nutrisyon at paghinga para sa lahat ng iba pang nabubuhay na organismo. Ang mga halaman ay mayroon ding kakayahan sa pagbabagong-buhay, maaari nilang ibalik ang mga vegetative organ.Ang agham na nag-aaral sa istraktura at buhay ng mga halaman, ang kanilang sistematiko, ekolohiya at pamamahagi ay tinatawag botanika(mula sa Greek botane - damo, halaman at logo - pagtuturo).Ang mga halaman ay bumubuo sa karamihan ng biosphere, na bumubuo sa berdeng takip ng Earth. Sila ay nakatira sa iba't ibang kondisyon- tubig, lupa, lupa-hangin na kapaligiran, sumasakop sa lahat ng lupain ng ating planeta, maliban sa mga nagyeyelong disyerto ng Arctic at Antarctica. uri ng buhay halaman. Ang mga puno ay nailalarawan sa pagkakaroon ng isang lignified stem - isang puno na nagpapatuloy sa buong buhay. Ang mga palumpong ay may ilang maliliit na tangkay. Ang mga damo ay nailalarawan sa pamamagitan ng makatas, berde, hindi lignified na mga shoots. Haba ng buhay. Mayroong taunang, biennial, pangmatagalang halaman. Ang mga puno at shrub ay mga perennial, habang ang mga halamang gamot ay maaaring mga perennial, annuals, o biennials. Istraktura ng halaman. Ang katawan ng halaman ay karaniwang nahahati sa ugat at ang pagtakas. Sa mas matataas na halaman, ang mga namumulaklak na halaman ang pinaka-organisado, marami at laganap. Bilang karagdagan sa ugat at shoot, mayroon silang mga bulaklak at prutas - mga organo na wala sa ibang mga grupo ng mga halaman. Maginhawang isaalang-alang ang istraktura ng mga halaman sa halimbawa ng mga namumulaklak na halaman. Ang mga vegetative organ ng mga halaman, ugat at shoot, ay nagbibigay ng kanilang nutrisyon, paglaki at asexual reproduction.
Mga uri ng root system: 1 - baras; 2 - mahibla; 3 - hugis-kono na ugat ng perehil; 4 - mga ugat ng beet ng sibuyas; 5 - dahlia root cones
Sa tulong ng () ugat, ang halaman ay naayos sa lupa. Nagbibigay din ito ng tubig at mineral, at kadalasang nagsisilbing site para sa synthesis at imbakan ng mga sustansya. Nagsisimulang mabuo ang mga ugat sa embryo ng halaman. Sa panahon ng pagtubo ng buto, nagbubunga ang germinal root pangunahing ugat. Pagkaraan ng ilang sandali, marami mga ugat sa gilid. Sa isang bilang ng mga halaman, nabubuo ang mga tangkay at dahon adventitious na mga ugat. Ang hanay ng lahat ng mga ugat ay tinatawag sistema ng ugat. Ang root system ay maaaring pamalo, na may mahusay na binuo pangunahing ugat (dandelion, labanos, puno ng mansanas) o mahibla, nabuo sa pamamagitan ng lateral at adventitious roots (barley, trigo, sibuyas). Ang pangunahing ugat sa naturang mga sistema ay hindi maganda ang binuo o ganap na wala. Sa isang bilang ng mga halaman, ang mga sustansya (almirol, asukal) ay nakaimbak sa mga ugat, halimbawa, sa mga karot, singkamas, at beet. Ang ganitong mga pagbabago ng pangunahing ugat ay tinatawag ugat pananim. Sa dahlias, ang mga sustansya ay idineposito sa makapal na mga ugat, tinatawag sila ugat tubers. Mayroong iba pang mga pagbabago sa mga ugat sa kalikasan: mga ugat ng attachment(sa baging, galamay-amo), mga ugat sa himpapawid(para sa monstera, orchid), nakapirming ugat(sa mga halaman ng bakawan - banyan), mga ugat ng paghinga(sa mga halaman ng latian). Ang ugat ay lumalaki sa tuktok kung saan matatagpuan ang mga selula pang-edukasyon na tisyu - isang punto ng paglago. Protektado siya takip ng ugat. mga buhok sa ugat sumipsip ng tubig na may mga mineral na natunaw dito suction zone. Sa pamamagitan ng sistema ng pagsasagawa tubig ugat at mineral umakyat sa mga tangkay at dahon, at ang mga organikong bagay ay gumagalaw pababa.
Ang pagtakas- Ito ay isang kumplikadong vegetative organ, na binubuo ng mga buds, stem at dahon. Kasama ng mga vegetative shoots, ang mga namumulaklak na halaman ay may mga generative shoots kung saan nabubuo ang mga bulaklak.Ang shoot ay nabuo mula sa germinal bud ng buto. Ang pagbuo ng mga perennial shoots mula sa mga buds ay malinaw na nakikita sa tagsibol. Ayon sa lokasyon ng bato sa tangkay, nakikilala nila apikal at mga lateral na bato. Tinitiyak ng apikal na usbong ang paglaki ng shoot sa haba, at ang mga lateral - ang sumasanga nito. Ang bato ay natatakpan sa labas na may mga siksik na kaliskis, madalas na pinapagbinhi ng mga resinous na sangkap, sa loob ay mayroong isang panimulang shoot na may isang kono ng paglago at mga leaflet. Sa mga axils ng mga paunang dahon, may mga halos hindi kapansin-pansin na mga panimulang putot. Sa generative bud ay ang mga simulain ng mga bulaklak.
stem- ito ang axial na bahagi ng shoot, kung saan matatagpuan ang mga dahon at mga putot. Gumaganap ito ng isang sumusuportang function sa halaman, tinitiyak ang paggalaw ng tubig at mineral mula sa ugat hanggang sa mga dahon, mga organikong sangkap - pababa, mula sa mga dahon hanggang sa ugat.Sa panlabas, ang mga tangkay ay magkakaiba: sa mais, mirasol, birch - patayo; sa wheatgrass, cinquefoil - gumagapang; sa bindweed, hops - kulot; sa mga gisantes, creepers, ubas - pag-akyat. Panloob na istraktura iba ang stem sa monocot at dicot na halaman ().
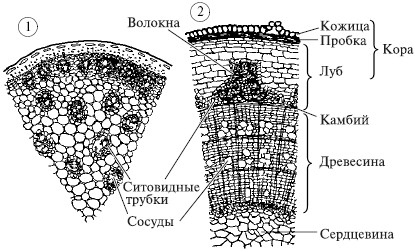
Panloob na istraktura ng tangkay. cross section: 1 - tangkay ng mais (ang mga vascular bundle ay matatagpuan sa buong tangkay); 2 - mga sanga ng linden
1. Sa halamang dicotyledonous ang tangkay ay natatakpan ng balat sa labas - epidermis, sa pangmatagalan lignified stems, ang balat ay pinalitan tapon. Sa ilalim ng cork ay isang bast na nabuo sa pamamagitan ng sieve tubes na tinitiyak ang paggalaw ng mga organikong sangkap sa kahabaan ng tangkay. Ang mga bast mechanical fibers ay nagbibigay ng lakas ng tangkay. Cork at bast form tumahol. Sa gitna ng bast ay cambium- isang solong layer ng mga cell ng pang-edukasyon na tisyu, na nagsisiguro sa paglaki ng stem sa kapal. Sa ibaba nito kahoy may mga sisidlan at mekanikal na hibla. Ang tubig at mga mineral na asing-gamot ay gumagalaw sa mga sisidlan, at ang mga hibla ay nagbibigay ng lakas sa kahoy. Habang lumalaki ang kahoy, nabubuo ito taunang singsing, kung saan natutukoy ang edad ng puno. Matatagpuan sa gitna ng tangkay core. Gumaganap ito ng isang function ng imbakan, ang organikong bagay ay idineposito dito. 2. Sa halamang monocot ang tangkay ay hindi nahahati sa bark, wood at pith; kulang sila ng cambial ring. Ang mga konduktibong bundle, na binubuo ng mga sisidlan at mga tubo ng salaan, ay pantay na ipinamamahagi sa buong tangkay. Halimbawa, sa mga cereal, ang tangkay ay isang dayami, guwang sa loob, at ang mga vascular bundle ay matatagpuan sa kahabaan ng periphery. Ang isang bilang ng mga halaman ay may binagong mga tangkay: mga tinik sa hawthorn, na nagsisilbing proteksyon; tendrils ng ubas - para sa paglakip sa isang suporta.
Sheet- Ito ay isang mahalagang vegetative organ ng isang halaman na gumaganap ng mga pangunahing function: photosynthesis, water evaporation at gas exchange.Mayroong ilang mga uri ng pag-aayos ng dahon sa mga halaman: susunod, kapag ang mga dahon ay salit-salit na nakaayos, kabaligtaran-dahon ay magkatapat whorled- tatlo o higit pang mga sheet ang umaalis sa isang node ().
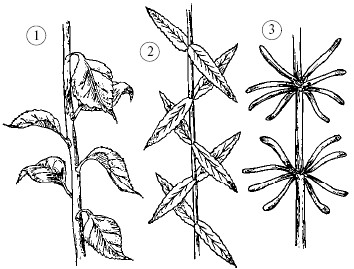
Pag-aayos ng dahon: 1 - regular; 2 - kabaligtaran; 3 - whorled
Ang sheet ay binubuo ng talim ng dahon at tangkay, minsan may mga stipule. Ang mga dahon na walang tangkay ay tinatawag laging nakaupo. Sa ilang mga halaman (cereal), ang mga dahon na walang tangkay ay bumubuo ng isang tubo - isang kaluban na bumabalot sa tangkay. Ang mga naturang dahon ay tinatawag vaginal().
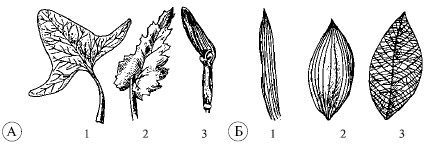
Mga uri ng dahon (A): 1 - petiolate; 2 - laging nakaupo; 3 - vaginal; leaf venation (B): 1 - parallel; 2 - arko; 3 - mesh
Ang mga dahon ay maaaring simple o tambalan. simpleng sheet ay may isang talim ng dahon, at magulo- ilang mga talim ng dahon na matatagpuan sa isang tangkay ().
Ang mga dahon ay simple: 1 - linear; 2 - lanceolate; 3 - elliptical; 4 - ovoid; 5 - hugis puso; 6 - bilugan; 7 - swept; kumplikado: 8 - ipinares; 9 - hindi ipinares; 10 - trifoliate; 11 - palad
Iba't ibang anyo ng mga talim ng dahon. Sa mga simpleng dahon, ang mga talim ng dahon ay maaaring buo at dissected na may iba't ibang mga gilid: may ngipin, may ngipin, crenate, kulot. Ang mga compound na dahon ay maaaring ipares at hindi magkapares na pinnate, palmate, trifoliate. May sistema sa leaf plate ugat, gumaganap ng pagsuporta at transportasyon function. Mayroong mesh venation (sa karamihan ng mga dicotyledonous na halaman), parallel (cereal, sedges) at arc (lily of the valley) (tingnan). Ang panloob na istraktura ng sheet (). Ang labas ng sheet ay natatakpan epidermis – balat, na pinoprotektahan ang mga panloob na bahagi ng dahon, kinokontrol ang palitan ng gas at pagsingaw ng tubig. Ang mga selula ng balat ay walang kulay. Sa ibabaw ng dahon ay maaaring may mga paglabas ng mga selula ng balat sa anyo ng mga buhok. Magkaiba ang kanilang mga tungkulin. Pinoprotektahan ng ilan ang halaman mula sa pagkain ng mga hayop, ang iba mula sa sobrang init. Ang mga dahon ng ilang halaman ay natatakpan ng wax coating na hindi pinapayagan ang kahalumigmigan na dumaan. Nakakatulong ito upang mabawasan ang pagkawala ng tubig mula sa ibabaw ng mga dahon.
Ang panloob na istraktura ng dahon: 1 - balat; 2 - stomata; 3 - columnar tissue; 4 - spongy tissue; 5 - ugat ng dahon
Sa ilalim ng dahon sa karamihan ng mga halaman sa epidermis ay marami stomata- mga butas na nabuo ng dalawang guard cell. Sa pamamagitan ng mga ito, ang pagpapalitan ng gas, pagsingaw ng tubig ay isinasagawa. Ang pagbubukas ng stomata ay bukas sa araw at sarado sa gabi. Ang panloob na bahagi ng sheet ay nabuo sa pamamagitan ng pangunahing asimilating tissue sumusuporta sa proseso ng photosynthesis. Binubuo ito ng dalawang uri ng berdeng selula - kolumnar, matatagpuan patayo, at bilugan, maluwag na matatagpuan espongha. Naglalaman ang mga ito ng isang malaking bilang ng mga chloroplast, na nagbibigay kulay berde sheet. Ang laman ng dahon ay natatakpan ng mga ugat na nabuo ng mga conductive vessel at sieve tubes, pati na rin ang mga hibla na nagbibigay ng lakas. Ang mga organikong sangkap na na-synthesize sa dahon ay gumagalaw sa mga ugat patungo sa tangkay at mga ugat, at ang tubig at mga mineral ay dumadaloy pabalik. Sa ating mga latitude, ang malawakang paglalagas ng mga dahon ay sinusunod taun-taon - nahulog ang dahon. Ang hindi pangkaraniwang bagay na ito ay may mahalagang adaptive na halaga, pinoprotektahan nito ang halaman mula sa pagkatuyo, pagyeyelo, at pinipigilan ang mga sanga ng mga puno na masira. Bilang karagdagan, sa mga patay na dahon, ang halaman ay napalaya mula sa mga sangkap na hindi kailangan at nakakapinsala dito. Maraming mga halaman ang may binagong mga dahon na gumaganap ng mga tiyak na function. Ang mga tendrils ng gisantes, na nakakapit sa isang suporta, ay sumusuporta sa tangkay, ang mga sustansya ay naka-imbak sa mga scaly na dahon ng sibuyas, pinoprotektahan ito ng mga barberry spines mula sa pagkain, ang mga sundew traps ay nakakaakit at nakakahuli ng mga insekto. Karamihan sa mga perennial herbaceous na halaman ay mayroon pagbabago sa pagtakas, na inangkop upang maisagawa ang iba't ibang mga function ().
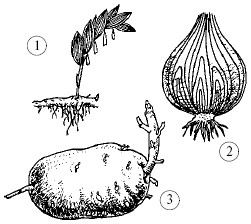
Mga pagbabago sa mga shoots: 1 - binili ng rhizome; 2 - bombilya ng sibuyas; 3 - patatas tuber
Rhizome- Ito ay isang binagong underground shoot na gumaganap ng mga function ng isang ugat, at nagsisilbi din upang mag-imbak ng mga nutrients at vegetative propagation ng mga halaman. Hindi tulad ng ugat, ang rhizome ay may kaliskis - binagong mga dahon at mga putot, ito ay lumalaki nang pahalang sa lupa. Lumalaki ang mga ugat mula rito. Ang rhizome ay matatagpuan sa lily of the valley, sedge, kupena, at couch grass. Ang mga strawberry ay bumubuo sa itaas ng lupa na binagong mga stolon - bigote na nagbibigay ng vegetative reproduction. Kapag nakikipag-ugnay sa lupa, nag-ugat sila sa tulong ng mga adventitious na ugat at bumubuo ng isang rosette ng mga dahon. Ang mga stolon sa ilalim ng lupa - mga tubers ng patatas - ay binagong mga shoots din. Ang mga sustansya ay naka-imbak sa isang mahusay na binuo core ng kanilang malakas na thickened stem. Sa mga tubers, maaari mong makita ang mga mata - mga buds na nakaayos sa isang spiral, kung saan nabuo ang mga shoots sa itaas ng lupa. Ang bombilya ay isang maikling shoot na may makatas na dahon. Ang mas mababang bahagi - ang ibaba ay isang pinaikling tangkay, kung saan lumalaki ang mga adventitious na ugat. Ang bombilya ay nabuo sa maraming mga liryo (mga tulip, liryo, daffodils). Ang mga binagong shoots ay ginagamit para sa vegetative propagation ng mga halaman.
Ang paggamit ng mga halaman para sa mga layuning panggamot ay umiral, ligtas nating masasabi, halos mula pa noong lumitaw ang sangkatauhan sa lupa. Ang halimbawa ng mga hayop ay malinaw na nagpapakita sa atin na sa mundo ng mga nabubuhay na nilalang ay may likas na konsepto kung ano ang pakinabang na maidudulot ng kaharian ng gulay. Ang tao, sa kanyang mausisa na pag-iisip, ay matagal nang naghangad na mapasok ang mga lihim at kababalaghan ng kahariang ito, at ang mga pangmatagalang obserbasyon at mga eksperimento ay nagturo sa kanya na gumamit ng mga halaman sa kanyang kalamangan. Sa bawat bansa, bago ang pag-unlad ng medisina bilang isang agham, may mga tao na mahusay na gumamit ng mga halaman para sa iba't ibang mga layuning panggamot, at ang kanilang karanasan ay mahusay na ginamit ng medikal na agham, na sa karamihan ng mga kaso ay sumasama sa kaharian ng gulay. Ang "Complete Russian Illustrated Dictionary-Herbal and Flower Garden" na inaalok sa atensyon ng mga mambabasa ay ang aming bagong tanyag na publikasyon, na nilayon namin na maglingkod para sa kapakinabangan ng bawat pamilyang Ruso at naglalayong ipakilala ang mambabasa sa mundo ng mga halaman. Ang aklat na ito ay may mga sumusunod na layunin: 1) ito ay dapat magsilbi bilang isang kinakailangang tulong para sa bawat magsasaka, na nagbibigay sa kanya ng mahalagang gabay sa paglilinang at paggamit ng ilang mga halaman sa kanyang sambahayan; 2) dapat itong magsilbi bilang isang desktop reference guide para sa mga doktor, paramedic, parmasyutiko, industriyalista at, sa pangkalahatan, lahat ng tao na gumagawa at nangangalakal ng mga produktong parmasyutiko; 3) pangunahing nilalayon niyang ipaalam sa bawat pamilyang Ruso ang mga pamamaraan ng paggamit ng kaharian ng halaman, kapwa para sa paggamit ng sambahayan at, lalo na, para sa mga layuning medikal - upang maibsan at pagalingin ang mga sakit sa tulong ng mga simpleng hindi nakakapinsalang mga remedyo na dinadala sa atin ng mga puno, bulaklak. , damo, ugat, atbp.
Aklat:
Sa paglalahad dito ng paglalarawan ng mga organo ng halaman, itinuring namin na kinakailangan, para sa higit na kalinawan, na ilarawan ito sa pamamagitan ng mga guhit, na sinabi ni Messrs. makikita ng mga mambabasa sa simula ng aklat, sa mga pahinang may pamagat na: "Mga Organo ng Halaman",
Ang isang buto o halamang namumulaklak ay karaniwang may mga vegetative na organo (ugat, tangkay, dahon), na binanggit sa itaas, at mga organo ng reproduktibo (bulaklak, prutas at buto).
Sa una, ang seed plant ay binubuo lamang ng mga vegetative organ; minsan (halimbawa, sa mga puno) lumipas ang ilang taon bago lumitaw ang mga organo ng reproduktibo, ibig sabihin, ang halaman ay namumulaklak.
Bumaling tayo sa paglalarawan ng mga vegetative organ, o mga organo ng nutrisyon, na maaaring bawasan sa apat na pangunahing mga: ugat, tangkay, dahon, buhok.
ugat. Ang simula ng ugat sa anyo ng ugat ay nasa binhi na. Sa panahon ng pagsibol, ang ugat na ito ay laging lumalago, lumalalim sa lupa, nagsasanga at nagiging ugat doon.
Ang ugat ay may dalawahang layunin: 1) ito ay nagsisilbing palakasin ang halaman at 2) ito ay kumukuha ng tubig at mga solusyon ng mga mineral na asin mula sa lupa, na nagsisilbing pagkain para sa halaman.
Alinsunod sa dalawahang layunin na ito ay ang lahat ng panlabas at panloob na mga katangian nito.
Ang ugat ay naiiba sa tangkay sa mga sumusunod na katangian: 1) palagi itong lumalaki pababa. Ito ay karaniwang ari-arian mga ugat, bagaman ito ay nagpapakita lamang ng sarili sa mga pangunahing ugat, habang ang kanilang pangalawang mga sanga ay maaaring magkaroon ng anumang direksyon; 2) sa ugat ay hindi natin makikita ang mga dahon at anumang katulad na pormasyon; 3) ang ugat ay hindi kailanman nagpapakita ng mga node; at 4) ang ugat ay hindi direktang gumagawa ng mga tangkay, ngunit tanging mga ugat.
Bilang karagdagan, ang mga ugat ay hindi humahaba sa kanilang buong haba, ngunit sa kanilang mga dulo lamang. Ang kanilang "punto ng paglago", ibig sabihin, ang pangkat ng mga batang selula na, sa pamamagitan ng kanilang pagpaparami, ay gumagawa ng isang pagpahaba ng ugat, ay hindi matatagpuan sa pinakadulo ng mga sanga ng ugat, ngunit natatakpan ng isang espesyal na takip na gawa sa luma, namamatay na tissue mula sa labas, na bumubuo ng "root cap". Ang mga batang bunga ng mga ugat na malapit sa kanilang mga dulo ay kadalasang natatakpan ng masaganang, pinong mga buhok, na tinatawag na "mga ugat ng buhok," na nagsisilbing pagkuha ng tubig at mga mineral na solusyon sa asin mula sa lupa. Mayroon ding mga purong anatomical na katangian na nagpapakilala sa ugat mula sa tangkay.
Sa hitsura, nakikilala nila ang "pangunahing ugat", iyon ay, ang nakuha mula sa pag-unlad ng ugat ng embryo at bumubuo, parang, isang direktang pagpapatuloy ng stem, mula sa "lateral o adventitious roots", nauugnay sa una, tulad ng mga sanga sa pangunahing tangkay.
Ang pangunahing ugat ay tinatawag na "branched" (Fig. 1) kung ito ay nahahati sa malalaking sanga, at "simple" (Fig. 2) kung ang mga ugat na umaabot mula dito ay medyo maliit.
Minsan ang pangunahing ugat ay nagiging napaka-mataba, cylindrical, hugis spindle o hugis singkamas, tulad ng mga karot, beets at singkamas. Gayunpaman, hindi laging posible na makilala ang pangunahing ugat. Sa maraming mga halaman, halimbawa, sa rye, trigo, atbp., ang ugat ng embryo ay nagyeyelo sa pinakadulo simula, at ang mga pangalawang ugat lamang ang bubuo. Ang buong ugat pagkatapos ay binubuo ng isang bundle ng mahaba, simple o branched roots; ang nasabing ugat ay tinatawag na "fibrous" (Fig. 3).
Minsan ang ilan sa mga buds, iyon ay, mga indibidwal na ugat, ay puno ng mga sustansya at lumalaki nang malakas sa kapal, na bumubuo ng mga tubers (Larawan 4).
Ang nasabing ugat ay tinatawag na "bumpy". Ayon sa pag-asa sa buhay ng mga ugat ay taunang, biennial at pangmatagalan. Sa mga annuals at biennials, ang ugat ay namamatay kasama ang pagpapatayo ng buong halaman, sa mga perennials, ang ugat ay nagiging makahoy at umiiral nang maraming taon sa parehong paraan tulad ng buong halaman. Ngunit sa mga pangmatagalang halaman na mala-damo, ang pangunahing ugat ay maaaring mamatay bago ang kamatayan ng halaman mismo at mapalitan ng mga bagong adventitious na ugat. Sa fibrous at knobby roots, ang mga indibidwal na lobes at tubers ay unti-unting namamatay, na pinalitan ng mga bago, nang hindi nakakagambala sa buhay ng buong root system.
Ang mga batang ugat ay nabuo hindi lamang sa mga ugat, ngunit maaari ring bumuo kahit saan sa tangkay. Sa mga halaman na may gumagapang na mga tangkay, ang pagbuo ng mga ugat sa tangkay ay madalas na sinusunod, pangunahin sa mga node nito. Ang mga nakaugat na bahagi ng tangkay ay pinaghihiwalay at maaaring humantong sa isang malayang buhay. Ang mga tuwid na tangkay ay hindi karaniwang nag-uugat, ngunit kung sila ay nadikit sa mamasa-masa na lupa (sa pamamagitan ng pagyuko ng mga sanga sa lupa, o sa pamamagitan ng paglakip ng isang collapsible na palayok ng mamasa-masa na lupa sa paligid ng sanga), sila, pagkaraan ng ilang oras, ay bubuo ng mga ugat. sa angkop na lugar. Ito ay ginagamit sa paghahardin para sa "layering".
stem. Ang tangkay ay isang organ na tumutubo sa tuktok at namumunga ng mga dahon. Nakatago pa rin ang rudiment nito sa mikrobyo ng binhi; kapag ang huli ay tumubo, ang batang tangkay ay nagsisimulang tumubo paitaas habang ang ugat ay dumadaloy pababa. This property to grow up is katangian stem sa pangkalahatan, bagaman sa partikular ay may mga pagbubukod sa panuntunang ito.
Bilang karagdagan sa pag-aari sa itaas, ang tangkay ay naiiba sa ugat dahil ito ay namumunga ng mga dahon at maaaring magbunga ng mga bagong tangkay at ugat; pagkatapos ay madalas itong nagpapakita ng mga buhol sa haba nito, tulad ng sa trigo, rye, at iba pa. (Larawan 5).
Ang tangkay ay lumalaki hindi lamang sa dulo nito, ngunit nagpapahaba sa lahat ng mga batang bahagi; ang punto ng paglago nito ay nasa pinakadulo at walang takip, ngunit natatakpan ng alinman sa mga batang dahon o ganap na hubad.
Sa hitsura, istraktura at pag-asa sa buhay, ang mga tangkay ay magkakaiba. Sa mga puno, sila ay nagiging napakatigas at nabubuhay ng maraming daan at libu-libong taon. Ang mga halamang damo ay may habang-buhay na limitado sa ilang buwan.
Ang ilang mga halaman ay may mga tangkay na napakaikli na tinatawag silang walang tangkay, habang ang iba ay hanggang ilang daang metro ang haba. Ang normal na posisyon ng tangkay ay maaaring ituring na patayo, na kadalasang matatagpuan sa iba't ibang uri ng mga halaman. Gayunpaman, hindi ito palaging pinapanatili. Sa ilang mga halaman, ang tangkay ay lumalaki pataas, ngunit masyadong mahina upang mapanatili ang isang tuwid na posisyon sa sarili nitong.
Ang mga naturang halaman ay may tangkay na maaaring umakyat (Larawan 6) o kumakapit sa mga dayuhang bagay na may mga tendrils, espesyal na attachment, curving petioles, atbp. Ang ganitong mga tangkay ay tinatawag na climbing. Sa iba pang mga halaman, ang tangkay ay direktang kumakalat sa lupa at sa parehong oras ay madalas na nag-ugat. Ang nasabing tangkay ay tinatawag na gumagapang (Larawan 7).

Tulad ng nabanggit sa itaas, sa mga pangmatagalang damo, ang bahagi na nananatili sa lupa ay nagpapalipas ng taglamig at sa tagsibol ay muling bumubuhay sa tangkay, na namamatay sa taglagas.
Ang mga underground na bahagi ng tangkay ay matatagpuan sa iba't ibang halaman magkaibang pinanggalingan, istraktura, hitsura at samakatuwid ay may iba't ibang mga pangalan. Sa pangkalahatan, ang mga ito ay:
Rhizome, ibig sabihin, isang mas marami o hindi gaanong pahabang bahagi ng tangkay, lumalaki nang pahalang o pahilig sa ilalim ng lupa (Larawan 8, 9).
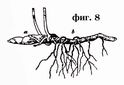
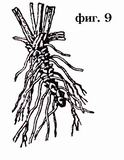
Ang pag-unlad ng rhizome ay pinakamadaling magawa sa paraang sa isang halaman na lumago mula sa isang buto, ang pangunahing ugat at ang base ng tangkay, na nasa lupa, ay nagpapanatili ng sigla nito sa panahon ng taglamig. Sa tagsibol, ang bahaging ito ng tangkay ay bubuo ng isang namumulaklak na tangkay mula sa lateral o apikal na usbong, na namamatay sa taglagas. Unti-unting namamatay, pagkatapos ng ilang taon, at ang pangunahing ugat, ang parehong bahagi ng tangkay ay patuloy na nabubuhay, patuloy na namamatay mula sa isang dulo at umuunlad mula sa kabilang dulo. Sa ganitong paraan, ang mga vertical o obliquely na lumalagong rhizomes ay nabuo. Ang mga ito ay kadalasang medyo maikli at may tuldok na mga ugat. Maaari silang makilala mula sa mga ugat sa pamamagitan ng mga bakas ng attachment ng mga dahon, sa anyo ng mga scars o nabubulok na petioles. Sa ibang mga kaso, ang underground na bahagi ng stem ay nagsisimulang lumaki nang nakapag-iisa sa ilalim ng lupa sa pahalang na direksyon. Ang ganitong mga rhizome ay maaaring maabot kung minsan ng napakahabang haba, tulad ng sa mga sedge, tambo, atbp. Sa kanila, maaari mong palaging makahanap ng mga paunang dahon sa anyo ng maliliit na kaliskis, ngunit sa mga halaman na may buhol na tangkay, ang rhizome ay palaging nagpapakita ng malinaw na mga node. Ang mga namumulaklak na tangkay ng naturang rhizome ay nabuo sa dalawang paraan: 1) alinman sa dulo ng rhizome ay nagbabago sa direksyon ng paglaki nito sa tagsibol, yumuko paitaas at lumalaki sa isang namumulaklak na tangkay, ang rhizome ay nagpapatuloy sa isang gilid na shoot na patuloy na lumalaki. pahalang; o, sa kabaligtaran, ang rhizome ay patuloy na humahaba sa isang pahalang na direksyon, at ang mga namumulaklak na tangkay ay lumilitaw bilang mga side shoots sa mga punto ng attachment ng mga dahon.
Ang una ay tinatawag na "panghuling" (tiyak) na mga rhizome, ang huli ay "walang katapusan" (hindi tiyak).
Ang mga underground shoots ay malapit sa uri ng rhizomes. Naiiba sila dahil mas mabilis silang lumalaki kaysa sa mga rhizome at may maikling pag-iral. Tumatakbo sa isang tiyak na distansya sa ilalim ng lupa, kadalasan ay lumalapit sila sa ibabaw ng lupa at dito nagkakaroon ng rosette ng mga dahon o nagpapakapal lamang sa dulo, na nagdedeposito ng mga sustansya dito.
Pagkatapos ang shoot mismo ay namatay sa buong haba nito, habang ang dulo lamang nito ay hibernate, na bumubuo ng isang namumulaklak na tangkay sa susunod na tag-araw, pati na rin ang mga bagong shoots.
Kasama rin sa uri ng naturang mga shoots ang mga sanga sa ilalim ng lupa ng tangkay, na sa mga patatas ay nagdadala ng mga tubers. Ang mga tubers na ito ay ang makapal na dulo ng mga shoots at nilagyan ng "mga mata", ibig sabihin, mga buds, kung saan ang mga tangkay sa itaas ng lupa ay bubuo.
Ang mga bombilya ay nagpapakita sa amin ng ibang uri (Larawan 11).
Ito rin ay isang underground wintering na bahagi ng stem; ngunit ang tangkay mismo ay napakaikli at kumakatawan sa tinatawag na "ibaba" ng bombilya; maraming kaliskis (i.e., binagong mga dahon) ang nakaupo dito, na bumubuo sa pangunahing masa ng bombilya. Nagdeposito sila ng maraming sustansya, na pagkatapos ay napupunta sa pagbuo ng isang namumulaklak na tangkay. Ang mga bombilya sa parehong paraan ay maaaring maibalik sa dalawang paraan: sa ilan, ang gitnang usbong ay bubuo sa isang namumulaklak na tangkay, at isang bagong bombilya ang bumubuo sa gilid; sa iba, ang bombilya ay patuloy na nabubuo mula sa panloob na usbong, at ang mga namumulaklak na tangkay ay nabuo mula sa mga lateral bud na umuunlad sa axil ng mga kaliskis.
Ang isang bagay sa pagitan ng isang rhizome at isang bombilya ay kinakatawan ng "tuber", halimbawa, sa safron. Sa hitsura, mukhang mga bombilya ang mga ito, ngunit hindi binubuo ng mga kaliskis, ngunit ng pamamaga ng tangkay. Ang kanilang pagpapanumbalik ay ginagawa sa parehong paraan tulad ng mga rhizome at bombilya.
Sa konklusyon, ang paglalarawan ng tangkay, sabihin natin tungkol sa anyo ng mga tangkay sa itaas ng lupa, na magkakaiba din, lalo na:
Ang hugis ng tangkay ay maaaring (sa seksyon) bilog, pipi, 3-4 o polyhedral. Ang mga tampok na ito ay madalas na pare-pareho para sa buong grupo: ang mga damo ay may mga bilog na tangkay, ang mga sedge ay trihedral, at ang mga labial ay tetrahedral.
Sheet. Ang mga dahon ay mga lateral na organo, kadalasang umuunlad sa anyo ng isang plato at nakaupo sa tangkay. Ang mga dahon ay naobserbahan na sa embryo ng buto, kung saan makikita natin ang mga ito sa dalawang anyo: 1) sa isang mas marami o hindi gaanong binuo na estado sa anyo ng mga cotyledon at 2) sa anyo ng primordia ng mga sumusunod na normal na dahon. Ang mga dahon ay naiiba sa mga ugat at tangkay sa mga sumusunod na katangian: 1) sa pamamagitan ng kanilang lateral na posisyon, 2) sa pamamagitan ng katotohanan na, habang palaging umuunlad sa tangkay, hindi na sila makakagawa ng anuman (ibig sabihin, hindi sila makakabuo ng anumang iba pang mga dahon, o mga tangkay. , o mga ugat), 3) paraan ng paglago.
Sa dahon, una sa lahat, nabuo ang tuktok nito, na bumubuo sa pinakalumang bahagi ng dahon, habang ang nakababata ay ang base nito. Ang sheet ay binubuo ng mga sumusunod na bahagi:
1) "mga plato", iyon ay, ang malawak, patag na bahagi nito; 2) ang "petiole" kung saan nakaupo ang plato, at 3) ang "sheath", iyon ay, ang extension ng petiole clasping ang stem, at sa wakas, ang "stipules" ay dapat ding maiugnay sa mga bahagi ng dahon (Fig . 12). Ito ay mga pormasyon na hugis dahon na nakaupo nang magkapares sa pagbuo ng dahon. Gayunpaman, hindi lahat ng mga bahaging ito ay kinakailangang naroroon. May mga dahon na walang tangkay, direktang nakaupo sa tangkay; ang mga naturang dahon ay tinatawag na "sessile". May mga dahon na may tangkay, ngunit walang kaluban, o may kaluban, ngunit walang tangkay (Larawan 13), o, sa wakas, walang tangkay at kaluban.
Ang mga Stipule ay wala din sa ilang mga halaman, habang sa iba ay patuloy silang sinusunod (Larawan 14); sabay sila iba't ibang uri, pagkatapos ay hugis-dahon, tulad ng sa mga gisantes (Larawan 15), pagkatapos ay lumaki nang magkasama sa anyo ng isang maikling tubo ("mga kampanilya"), tulad ng sa bakwit (Larawan 16), o maging mga spine, tulad ng sa mga gooseberry (Fig . 17).


Sa wakas, kahit na pangunahing bahagi dahon, lalo na ang plato, ay maaaring wala at mapalitan ng isang tangkay o stipules.
Ang talim ng dahon ay binubuo ng "leaf pulp" na tinusok ng "veins" o "nerves"; ang mga ugat na ito ay nagmumula sa tangkay, sumasanga sa iba't ibang direksyon at bumubuo, kumbaga, ang balangkas ng dahon. Ayon sa likas na katangian ng nerve (i.e., ayon sa paraan ng pagsasanga ng mga nerbiyos ng dahon), ang mga dahon ay naiiba sa "arc-nervous" at "co-nervous". Sa una, ang mga nerbiyos ay halos hindi sumanga; umuusbong mula sa mga base ng dahon, tumatakbo ang mga ito halos magkatulad, tulad ng sa mga cereal, o, kurbadong sa isang arcuate na paraan, muling nagsalubong sa tuktok ng dahon. Sa mga dahon ng conjunctural, ang mga lateral nerve ay umaalis sa mga anggulo mula sa isa o higit pang mga pangunahing nerbiyos, na, sa turn, ay muling sumanga.
Ang mga dahon ng arcuate ay pangunahing katangian ng mga monocotyledonous na halaman, habang ang mga angulated ay katangian ng mga dicotyledonous, ngunit sa partikular mayroon ding mga paglihis mula sa pangkalahatang tuntunin.
Ayon sa lokasyon ng mga nerbiyos, ang mga carbonervous na dahon ay nahahati sa: mga pinnate na dahon, kapag ang petiole ay nagpapatuloy sa isang pangunahing nerve, kung saan ang mga lateral ay umaabot sa lahat ng direksyon; palmar nerve (finger nerve), kapag ang tangkay sa pinaka-base ng dahon ay nahati sa ilang nerbiyos na magkapareho ang laki, na nag-iiba tulad ng mga daliri ng isang kamay (Larawan 18); thyroid nerve, kapag ang tangkay ay nagtatapos sa gitna ng plato, at ang mga nerbiyos, na halos pareho ang laki, ay naghihiwalay sa radii sa lahat ng direksyon (Fig. 19).
Ayon sa dismemberment ng plato, sila ay nakikilala: ang mga dahon ay buo, kung hindi sila kumakatawan sa anumang mga lobe at hiwa; lobed, kung ang mga ito ay nahiwa hanggang sa isang ikatlo o hanggang kalahati ng distansya sa pagitan ng margin at ang pangunahing nerve (o isa sa mga pangunahing nerbiyos), at kung, bukod dito, ang mga nakausli na bahagi ng dahon o vane ay maikli at mapurol ( igos 20-22); split, kung sila ay insed sa parehong lalim, ngunit ang kanilang mga lobes ay makitid at matalim; hiwalay kung ang plato ay nahahati halos sa pangunahing nerve (fig. 23 at 18), ngunit ang mga lobe ay konektado pa rin sa base ng pulp ng dahon; dissected kung sila ay nahahati sa nerbiyos, upang ang mga indibidwal na bahagi (na tinatawag na mga segment dito) ay konektado lamang sa pamamagitan ng mga sanga ng pangunahing nerve tulad ng isang tangkay (fig. 24-25).
Ang lahat ng mga hugis ng dahon na ito ay tinatawag na simple. Ang mga compound na dahon ay ang mga kung saan ang lamina ay nahahati sa ganap na magkakahiwalay na mga bahagi (tinatawag na mga leaflet sa kasong ito), nakaupo sa mga ramification ng tangkay at konektado sa mga huling espesyal na joints (Fig. 26-27). Sa bagay na ito, ang mga dahon ng tambalan ay naiiba sa mga pinaghiwa-hiwalay, kung saan sila ay madalas na may malaking pagkakahawig.
Kapag ang tambalang dahon ay namatay at nalalanta, tulad ng, halimbawa, sa puti at dilaw na akasya, ang bawat leaflet ay nahuhulog nang hiwalay mula sa karaniwang tangkay, habang ang isang dissected na dahon, na walang mga kasukasuan, ay bumagsak nang buo.
Ayon sa pangkalahatang balangkas ng dahon, ibig sabihin, hindi pinapansin ang mga hiwa ng plato, ang mga dahon ay nakikilala: bilog, na kumakatawan sa humigit-kumulang isang bilog (Larawan 28), hugis-itlog (Larawan 29), ovoid (Larawan 30), pahaba , elliptical, lanceolate (Fig. 31) at linear (Fig. 32).
Ang lahat ng mga terminong ito ay inilalapat din sa mga indibidwal na leaflet at leaf lobes. Ang mga dahon ay hugis-puso din (Larawan 33), hugis-kidney (Larawan 34), hugis-arrow (Larawan 35), hugis-sibat (Larawan 36), spatulate (Larawan 37).
Ang mga dahon ay tinatawag na stalk-bearing kung ikinakapit nila ang tangkay gamit ang kanilang base (Larawan 13), tinutusok kung ang kanilang mga gilid ay tumubo nang magkasama sa paligid ng tangkay (Larawan 38).
Ang lahat ng mga terminong ito ay mahalaga sa paglalarawan ng mga halaman, dahil ang hugis ng mga dahon ay isang napakahalagang katangian para sa kahulugan ng mga halaman.
Iba-iba ang pagkakaayos ng mga dahon sa tangkay. Ang mga dahon ay nakikilala sa pamamagitan ng pag-aayos ng dahon: variable (o kahalili), kung sila ay nakaupo sa tangkay nang isa-isa sa walang nakikitang pagkakasunud-sunod, kabaligtaran, kung sila ay nakaupo sa dalawa, isa laban sa isa (Fig. 44) at naka-ring o whorled, kung sila ay umupo. sa ilang mga singsing sa paligid ng tangkay (Larawan 45).
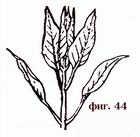

Bilang karagdagan, ang mga dahon ay nakikilala: ang mga katutubo, na matatagpuan sa ilalim ng lupa na bahagi ng tangkay, sa base nito, o palibutan ang mga buds ng makahoy na mga halaman, madalas silang walang berdeng plato at kumukuha ng anyo ng mga kaluban o kaliskis; apikal, na matatagpuan malapit sa mga bulaklak; iba ang anyo nila kaysa berde, minsan iba ang kulay.
Ang halaga ng mga dahon para sa mga halaman ay napakahalaga. Sila ay, tulad ng ugat at tangkay, ang mga nutritional organ ng halaman, na gumagawa ng ilang mga organikong sangkap, habang ang ugat ay nagbibigay ng tubig at mineral.
Antennae at spines. Ang mga spine at tendrils ay nabibilang sa mga espesyal na pagbabago ng mga dahon. Ang antennae ay mahahabang pormasyon na parang sinulid na lumilitaw sa tangkay o sa mga dahon ng ilang halaman. Sa una sila ay medyo tuwid, ngunit, sa pagpindot sa ilang mga banyagang katawan, nagsisimula silang umikot sa paligid nila at suportahan ang mahinang tangkay, na nagpapahintulot na umakyat ito nang mas mataas at mas mataas. Sa mga halaman na ang tangkay ay sapat na malakas upang tumayo sa sarili nitong, ang mga tendrils ay hindi matatagpuan.
Ang mga tendrils ay may iba't ibang pinagmulan, ngunit sa pangkalahatan ay kumakatawan sa alinman sa pagbabago ng tangkay o dahon. Sa mga gisantes, ang huling dahon ng isang pinnate na dahon, kung minsan ang buong pinnate na dahon, ay nagiging isang tendril. Minsan ang mga stipule ay nagiging tendrils, o ang sanga mismo ay nagiging tendril sa dulo, at ang stem ay nagpapatuloy sa isang side shoot, tulad ng isang ubas. Sa ilang mga halaman, ang mga tunay na tendrils ay hindi nabuo, ngunit ang kanilang papel ay ginagampanan ng pag-twist ng mga karaniwang petioles ng mga pinnate na dahon.
Ang mga spine ay pareho ang pinagmulan ng antennae. Katulad nito, ang huling dahon ng isang pinnate na dahon ay maaaring maging malambot na gulugod, o ang buong karaniwang tangkay ay nagiging gulugod. Ang mga stipule ay maaari ding maging mga tinik, tulad ng isang puting akasya, o, sa wakas, mga buong sanga, tulad ng isang blackthorn.
Ang mga tinik ay dapat na makilala mula sa mga tinik, na nakakalat sa mga sanga nang walang anumang pagkakasunud-sunod, tulad ng, halimbawa, sa isang rosas. Ang mga tinik ay mga bunga lamang ng panlabas na himaymay at walang kinalaman sa mga organo ng halaman.
Bud. Ang bato ay isang kumplikadong organ na binubuo ng isang pinaikling tangkay at maraming leaf primordia. Ito ang simula ng isang bagong pagtakas. Ang isang stem ay bubuo mula sa usbong ng embryo; sa dulo nito, nagtataglay ito ng apical bud, na nagsisilbi para sa karagdagang pag-unlad nito. Ang mga lateral na sanga ay bubuo mula sa axillary, ibig sabihin, na matatagpuan sa mga sulok ng mga dahon na nakakabit sa tangkay. Ito ang normal na posisyon ng mga bato.
Sa mala-damo na mga tangkay, ang mga putot na inilatag sa mga axils ay agad na nagsisimulang umunlad. Sa mga palumpong at puno, gayunpaman, ang mga buds na nabuo sa tag-araw ay nakasuot ng kayumangging parang balat na mga kaliskis, taglamig sa form na ito at nagsisimulang umunlad sa susunod na tagsibol (Fig. 40).
Ang mga bud ay nagsisilbi hindi lamang para sa pagbuo ng mga bagong shoots, ngunit kung minsan para sa pagpaparami ng mga bagong halaman. Ang ganitong mga buds ay karaniwang may mataba na dahon na may malaking suplay ng sustansya, kaya naman tinawag silang mga sibuyas. Habang ang mga normal na buds ay palaging nabuo sa ilang mga lugar, lalo na, pangunahin sa mga axils ng mga dahon, ang tinatawag na adnexal buds ay maaaring umunlad sa lahat ng dako: sa ugat, sa tangkay, at sa mga dahon. Ito ay isang medyo bihirang kababalaghan, ngunit ito ay ginagamit para sa pagpapalaganap ng halaman sa pamamagitan ng mga segment ng mga ugat, tulad ng sa ilang mga species ng puno, o mataba na dahon, tulad ng sa ilang mga begonias.
Mga buhok o trichomes. Ang mga buhok, isa sa mga vegetative organ ng isang halaman, ay mga labasan ng pinakalabas na layer ng mga selula, ang tinatawag na epidermis. Sa maraming mga halaman sila ay ganap na wala sa mga organo sa ilalim ng lupa, sa iba pa sila ay sagana at sumasakop sa lahat ng mga organo. Naghahain sila ng iba't ibang layunin: ang isang makapal na balbon na takip ay maaaring maprotektahan ang halaman mula sa malakas na paglamig o, sa kabaligtaran, mula sa malakas na pag-init ng araw. Pinoprotektahan ng matitigas na buhok ang halaman mula sa mga slug, habang ang mga nakakatusok na buhok, tulad ng mga nettle, ay mula rin sa mga baka.
Sa wakas, may mga glandular na buhok na naglalabas ng iba't ibang mahahalagang langis.
Ang mga halaman, tulad ng lahat ng nabubuhay na bagay, ay binubuo ng mga selula. Daan-daang mga cell na may parehong hugis at may parehong function ay bumubuo ng isang tissue; ang isang organ ay binubuo ng ilang mga tissue. Ang mga pangunahing organo ng halaman ay ang mga ugat, tangkay at dahon, bawat isa sa kanila ay gumaganap ng isang napaka tiyak na pag-andar. Ang mahahalagang organo ng reproduktibo ay mga bulaklak, prutas, at buto.
Mga ugat
Ang mga ugat ay may dalawang pangunahing tungkulin: ang una ay ang pagpapakain sa halaman, ang pangalawa ay ang pag-aayos nito sa lupa. Sa katunayan, ang mga ugat ay sumisipsip ng tubig at mga mineral na asing-gamot na natunaw dito mula sa lupa, sa gayon ay nagbibigay sila ng patuloy na supply ng kahalumigmigan sa halaman, na kinakailangan kapwa para sa kaligtasan nito at para sa paglago nito. Iyon ang dahilan kung bakit napakahalaga na ang halaman ay hindi nalalanta o natuyo, regular itong diligin sa mainit at tuyo na oras.
Ang panlabas na nakikitang bahagi ng ugat ay ang makinis, walang buhok na lumalagong bahagi kung saan nangyayari ang pinakamataas na paglaki. Ang lumalagong punto ay natatakpan ng isang manipis na proteksiyon na kaluban, ang takip ng ugat, na nagpapadali sa pagpasok ng ugat sa lupa. Ang suction zone, na matatagpuan malapit sa punto ng paglago, ay idinisenyo upang sumipsip ng tubig at mga mineral na asing-gamot na kailangan ng halaman, ito ay natatakpan ng makapal na pababa, na madaling makita ng isang magnifying glass at kung saan ay binubuo ng pinakamanipis na ugat na tinatawag na ugat. mga buhok. Ang conductive zone ng mga ugat ay gumaganap ng function ng paglilipat ng mga baterya. Bilang karagdagan, mayroon din silang function ng suporta, matatag nilang inaayos ang halaman sa lupa. Ang hugis, sukat, istraktura at iba pang mga katangian ng mga ugat ay malapit na nauugnay sa mga pag-andar na ito at, siyempre, nagbabago depende sa kapaligiran kung saan kailangan nilang bumuo. Kadalasan ang mga ugat ay nasa ilalim ng lupa, ngunit mayroong tubig at panghimpapawid.
Ang mga ugat, kahit na sa mga halaman ng parehong species, ay ibang-iba ang haba, na depende sa uri ng lupa at sa dami ng tubig na nilalaman nito. Sa anumang kaso, ang mga ugat ay mas mahaba kaysa sa iniisip natin, lalo na kung isasaalang-alang natin ang pinakamanipis na buhok ng ugat, na ang layunin ay sumipsip; sa pangkalahatan, ang root apparatus ay higit na binuo kaysa sa aerial na bahagi ng halaman na matatagpuan sa ibabaw ng lupa.
stem
Ang pangunahing pag-andar ng stem ay upang suportahan ang aerial na bahagi at ang koneksyon sa pagitan ng root system at mga dahon, habang ang stem ay kumokontrol. pare-parehong pamamahagi nutrients sa buong internal organs ng halaman. Sa tangkay, kung saan ang mga dahon ay nakakabit, kung minsan ay medyo kapansin-pansin na mga pampalapot, na tinatawag na mga node, ang bahagi ng tangkay sa pagitan ng dalawang node ay tinatawag na internode. Ang stem, depende sa density nito, ay may iba't ibang pangalan:
Ang tangkay, kung hindi masyadong siksik, tulad ng karamihan sa mga mala-damo na halaman;
Ang dayami, kung ito ay guwang at nahahati, tulad ng sa mga cereal, sa pamamagitan ng malinaw na nakikitang mga buhol. Kadalasan mayroong maraming silica sa naturang tangkay, na nagpapataas ng lakas nito;
Ang puno, kung makahoy at may sanga, tulad ng karamihan sa mga puno; o makahoy, ngunit hindi sanga, na may mga dahon sa tuktok, tulad ng mga palad.
Depende sa density ng stem, ang mga halaman ay nahahati sa:
Herbaceous, na may pinong, hindi lignified stem;
Semi-shrub, kung saan ang stem lignifies ang puno ng kahoy lamang sa base;
Shrub, kung saan ang lahat ng mga sanga ay lignified, sumasanga mula sa pinaka-base;
Woody, kung saan ang puno ng kahoy ay ganap na lignified, mayroon itong gitnang axis (ang puno ng kahoy mismo), sumasanga lamang sa itaas na bahagi.
Depende sa habang-buhay na nauugnay sa ikot ng buhay, ang mga mala-damo na halaman ay karaniwang inuri bilang mga sumusunod:
Annuals, o annuals, kung sila ay tumubo lamang ng isang taon, at mamatay pagkatapos na sila ay mamulaklak, magbunga, at magkalat ng mga buto;
Biennials, o biennials, kung sila ay lumalaki sa loob ng dalawang taon (karaniwan ay sa unang taon mayroon lamang silang isang rosette ng mga dahon, sa ikalawang taon ay namumulaklak, namumunga, pagkatapos ay natuyo);
Ang mga perennial, o mga perennial, kung sila ay nabubuhay nang higit sa dalawang taon, kadalasan ay namumulaklak at namumunga bawat taon, at "pahinga", iyon ay, sa malamig o tuyo na mga panahon, ang nasa itaas na bahagi ng halaman ay namamatay, ngunit ang ilalim ng lupa. bahagi ng halaman ay nananatiling buhay. May mga halaman kung saan ang bahagi ng tangkay ay maaaring magbago at maging isang tunay na organ ng imbakan. Kadalasan ang mga ito ay mga tangkay sa ilalim ng lupa na nagsisilbi para sa vegetative propagation, pati na rin para sa pangangalaga ng halaman sa isang hindi kanais-nais na panahon para sa paglago. Ang pinakasikat sa kanila ay mga tubers (tulad ng patatas), rhizomes (iris) at mga bombilya (narcissus, hyacinth, sibuyas).
Mga dahon
Ang mga dahon ay may maraming iba't ibang mga pag-andar, ang pangunahing isa ay ang nabanggit na photosynthesis, iyon ay, isang kemikal na reaksyon sa tisyu ng dahon, sa tulong kung saan hindi lamang mga organikong sangkap ang nilikha, kundi pati na rin ang oxygen, na kinakailangan para sa buhay sa ating planeta. . Karaniwan ang dahon ay binubuo ng isang tangkay, isang talim ng dahon na mas malawak o hindi gaanong lapad, na sinusuportahan ng mga ugat, at mga stipule. Ang tangkay ay nag-uugnay sa dahon sa tangkay. Kung walang petiole, kung gayon ang mga dahon ay tinatawag na sessile. Sa loob ng dahon ay may mga vascular fibrous bundle. Nagpapatuloy sila sa talim ng dahon, sumasanga, bumubuo ng isang siksik na network ng mga ugat (nervation), kung saan ang katas ng halaman ay nagpapalipat-lipat, bilang karagdagan, sinusuportahan nila ang talim, na nagbibigay ng lakas. Batay sa lokasyon ng mga pangunahing ugat, mayroong iba't ibang uri ng venation: palmate, pinnate, parallel at arcuate. Ang talim ng dahon, depende sa kung saang halaman kabilang ito, ay may iba't ibang densidad (matigas, makatas, atbp.) at ganap. iba't ibang anyo(bilog, elliptical, lanceolate, sagittate, atbp.). At ang gilid ng talim ng dahon ay nakakakuha ng pangalan nito depende sa istraktura nito (solid, serrated, serrated, lobed, atbp.). Kung ang bingaw ay umabot sa gitnang ugat, kung gayon ang mga lobe ay nagiging independyente at maaaring tumagal ng anyo ng mga leaflet, kung saan ang mga dahon ay tinatawag na kumplikado, sila naman, ay nahahati sa palmate-complex, pinnate-complex, at iba pa.
mga bulaklak
Ang kagandahan at pagka-orihinal ng mga hugis at kulay ng mga bulaklak ay may isang tiyak na layunin. Sa lahat ng ito, iyon ay, mga trick at device na binuo sa paglipas ng mga siglo, ang likas na katangian sa pana-panahon ay nagbibigay ng bulaklak lamang upang ang genus nito ay magpatuloy. Isang bulaklak na may lalaki at mga babaeng organo, upang makamit ang layuning ito, dapat itong dumaan sa dalawang pinakamahalaga at kinakailangang proseso: polinasyon at pagpapabunga. Karaniwan, sa mas mataas na mga halaman, ang mga bulaklak ay bisexual, iyon ay, mayroon silang parehong lalaki at babaeng organo. Sa ilang mga kaso lamang ang mga kasarian ay pinaghihiwalay: sa dioecious, halimbawa, sa willow, holly, at laurel, ang mga lalaki at babaeng bulaklak ay nasa iba't ibang mga specimen, at sa monoecious, halimbawa, sa mais at kalabasa, parehong lalaki at babae na mga bulaklak ay hiwalay na inilagay sa parehong halaman. Sa katunayan, ang lahat ng mga bahagi na bumubuo sa isang bulaklak ay iba't ibang mga pagbabago ng dahon na naganap upang maisagawa ang iba't ibang mga function.
Sa itaas ng peduncle, makikita mo ang isang pampalapot na tinatawag na receptacle, kung saan matatagpuan ang iba't ibang bahagi ng bulaklak. Ang doble, o simple, perianth ay ang panlabas at pinakakapansin-pansing bahagi ng bulaklak, ang perianth sa totoong kahulugan ng salita ay sumasaklaw sa mga organo ng reproduktibo at binubuo ng isang takupis at isang talutot. Ang calyx ay binubuo ng mga leaflet, kadalasang berde, tinatawag na sepals, ang kanilang gawain, lalo na sa panahon kung kailan ang bulaklak ay nasa yugto ng usbong, ay protektahan. panloob na mga bahagi. Kapag ang mga sepal ay pinagsama, tulad ng sa isang carnation, ang takupis ay tinatawag na sympetalous, at kapag sila ay pinaghiwalay, halimbawa, tulad ng sa isang rosas, ang takupis ay hiwalay na talulot. Ang calyx ay bihirang bumagsak, at sa ilang mga kaso hindi lamang ito nananatili, ngunit lumalaki din upang mas mahusay na matupad ang proteksiyon na function nito. Corolla - ang pangalawang elemento ng perianth - ay binubuo ng mga petals, karaniwang maliwanag na kulay at kung minsan ay kaaya-aya na amoy. Ang kanilang pangunahing pag-andar ay upang maakit ang mga insekto upang mapadali ang polinasyon at, nang naaayon, pagpaparami. Kapag ang mga petals ay higit pa o mas kaunting soldered magkasama, ang talutot ay tinatawag na cleavage, at kung sila ay hiwalay, pagkatapos ay hiwalay na talulot. Kapag walang malinaw na pagkakaiba sa pagitan ng calyx at corolla, tulad ng, halimbawa, sa isang tulip, ang perianth ay tinatawag na simpleng corolla, at ang bulaklak mismo ay simple. Ang reproductive male apparatus ng isang bulaklak, o androecium, ay binubuo ng isang variable na bilang ng mga stamen, na binubuo ng isang sterile, manipis at pahabang stamen na tinatawag na stamen filament, sa tuktok nito ay isang anther, naglalaman ito ng mga pollen sac. Ang pollen, ang nakakapataba na elemento ng lalaki, ay karaniwang dilaw o orange ang kulay.
Ang reproductive female apparatus ng isang bulaklak, o gynoecium, ay nabuo sa pamamagitan ng isa o higit pang mga pistil. Ang bawat isa sa kanila ay binubuo ng isang mas mababang guwang at namamaga na bahagi, na tinatawag na obaryo, na naglalaman ng isa o higit pang mga ovule, ang itaas na bahagi ng filiform ay tinatawag na haligi, at ang tuktok nito, na idinisenyo upang kolektahin at hawakan ang mga butil ng pollen, ay tinatawag na stigma.
Ang mga bulaklak sa isang halaman ay maaaring matatagpuan nang paisa-isa, sa tuktok o sa mga axils ng mga sanga, ngunit mas madalas silang pinagsama sa mga grupo, ang tinatawag na mga inflorescences.
Kabilang sa mga inflorescences, ang pinaka-karaniwan ay ang mga sumusunod: inflorescences na nabuo sa pamamagitan ng mga bulaklak sa pedicels: isang brush, halimbawa, wisteria, panicle (lilac), payong (karot) at kalasag, tulad ng isang peras. Ang mga inflorescences na nabuo sa pamamagitan ng walang stem, iyon ay, sessile na bulaklak: tainga (trigo), hikaw (hazel), basket (daisy).
polinasyon
Kadalasan, ang hangin, tubig, mga insekto at iba pang mga hayop ay nakikibahagi sa pinakamahalagang operasyon ng polinasyon, na kinakailangan para sa pagpaparami ng mga halaman. Maraming mga insekto, tulad ng mga bubuyog, bumblebee at butterflies, sa paghahanap ng nektar, isang matamis na sangkap na matatagpuan sa mga nectaries na matatagpuan sa panloob na bahagi ng maraming mga bulaklak, umupo sa mga bulaklak. Kapag hinawakan nila ang mga stamen, ang pollen mula sa mature anthers ay bumabagsak sa kanila, at inililipat nila ito sa iba pang mga bulaklak, kung saan ang pollen ay nahuhulog sa stigma. Ito ay kung paano nangyayari ang pagpapabunga. Ang maliwanag na kulay, kaakit-akit na hugis, at halimuyak ng mga bulaklak ay may isang tiyak na tungkulin ng pag-akit ng mga pollinating na insekto, na nagdadala ng pollen mula sa isang bulaklak patungo sa isa pa.
Ang pollen, lalo na ang napakagaan na pollen, na napakarami sa mga halaman na may maliliit na bulaklak na walang corolla, at samakatuwid ay hindi kaakit-akit sa mga insekto, ay dinadala din ng hangin. Ang pollen na ito, na dinadala sa maraming dami sa hangin, ang sanhi ng karamihan sa mga allergy sa tagsibol.
Mga prutas at buto
Pagkatapos ng pagpapabunga, ang mga dingding ng obaryo ay sumasailalim sa malalim na pagbabago, nagiging lignified o mataba, bumubuo sila ng isang prutas (o pericarp, testis), sa parehong oras, ang mga ovule ay bubuo. Nag-iipon ng isang supply ng nutrients, sila ay nagiging mga buto. Kadalasan, kapag hinog na ang prutas, ito ay malasa, mataba, matingkad ang kulay at mabango. Sa pamamagitan nito ay umaakit siya ng mga hayop, sa pamamagitan ng pagkain sa kanya, tinutulungan nila ang pagkalat ng mga buto. Kung ang prutas ay hindi maliwanag na kulay at hindi mataba, kung gayon ang mga buto nito ay magkakaibang kalat. Halimbawa, ang bunga ng meadow dandelion ay may magaan na fluffs, na kahawig ng isang maliit na parasyut, at ang mga bunga ng maple at linden ay may mga pakpak at madaling dinadala ng hangin; ang ibang mga prutas, gaya ng burdock, ay may mga kawit kung saan sila ay nakakapit sa lana ng tupa at sa damit ng tao.
Kabilang sa mga mataba na prutas, ang pinakasikat ay ang drupe, sa loob nito ay may isang buto na pinoprotektahan ng pericarp (cherry, plum, olive), at isang berry, kung saan karaniwang maraming buto at sila ay direktang inilubog sa pulp ( ubas, kamatis).
Ang mga tuyong prutas ay kadalasang inuuri sa bukas (cracking) at non-opening (non-cracking) depende sa kung sila ay bumuka sa kanilang sarili kapag hinog o hindi. Halimbawa, ang unang grupo ay kinabibilangan ng beans, o legume pods (peas, beans), leaflets (levkoy, labanos, beetroot), box (poppy) at achene (wrestler). Sa mga bunga ng pangalawang grupo ay palaging may isang buto, halos ibinebenta sa prutas mismo. Ang pinakasikat na mga halimbawa ay ang weevil sa mga cereal, ang lionfish sa maple at elm, at ang achene na may tufted sa Compositae.
Sa loob ng prutas ay isang buto kung saan mayroong isang embryo, halos isang hinaharap na halaman sa maliit na larawan. Sa sandaling nasa lupa, kung saan ang buto ay maaaring tumubo, ito ay umalis sa estado ng dormancy, kung saan kung minsan ay maaaring manatili kahit na ilang taon, at nagsisimulang umusbong. Kaya, nakumpleto ng binhi ang pag-andar nito, iyon ay, ang proteksyon at nutrisyon ng usbong, na hindi maaaring umiral nang nakapag-iisa, at nagsisimula ang isang bagong buhay.
,
,
Ang daloy ng tubig sa root system ay bumababa sa pagbaba ng temperatura. Nangyayari ito sa mga sumusunod na dahilan: 1) tumataas ang lagkit ng tubig, at samakatuwid ay bumababa ang mobility nito; 2) ang pagkamatagusin ng protoplasm para sa tubig ay bumababa; 3) ang paglago ng ugat ay pinipigilan; 4) bumababa ang rate ng metabolic process. Bumababa ang daloy ng tubig sa pagkasira ng aeration ng lupa. Ito ay mapapansin kapag, pagkatapos ng malakas na ulan, ang lupa ay binaha ng tubig, ngunit sa maliwanag na sikat ng araw, dahil sa malakas na pagsingaw, ang
nalalanta ang mga anino. Pinakamahalaga may konsentrasyon ng solusyon sa lupa. Ang tubig ay pumapasok lamang sa ugat kapag ang potensyal ng tubig ng ugat ay mas mababa kaysa sa potensyal ng tubig ng lupa. Kung ang solusyon sa lupa ay may mas negatibong potensyal, ang tubig ay hindi papasok sa ugat, ngunit lalabas dito.
Para sa normal na paggana, ang mga selula ng halaman ay dapat na puspos ng tubig. Ang estado ng saturation ay pinananatili sa tulong ng dalawang magkakaugnay na proseso: ang pag-agos at paglabas (paggasta) ng tubig, na bumubuo sa pagpapalitan ng tubig ng mga halaman. Ang ratio sa pagitan ng mga prosesong ito ay tinatawag na balanse ng tubig.
Ang halaman ay naglalabas ng tubig sa likido at singaw na estado. Ang proseso ng physiological ng pagsingaw ng tubig ng mga terrestrial na organo ng mga halaman, tulad ng nabanggit na, ay tinatawag na transpiration, ang pagpapalabas ng tubig sa isang drop-liquid state - guttation.
Nagmula ang buhay sa karagatan. Sa paglabas ng mga halaman sa lupa, may panganib ng kanilang kamatayan mula sa pagkatuyo. Bakit? Ang konsentrasyon ng singaw ng tubig sa hangin, kahit na sa isang mahalumigmig na klima, ay palaging mas mababa kaysa sa mga intercellular space ng dahon, samakatuwid, ang tubig ay hindi maiiwasang gumagalaw mula sa katawan ng halaman patungo sa kapaligiran, habang ang pagsasabog ng mga molekula ng tubig mula sa ibabaw ng dahon patungo sa hangin ay nangyayari nang 1,500 beses na mas mabilis kaysa sa kanilang pagpasok mula sa lupa patungo sa ugat.
Ang malaking pagkawala ng tubig ng halaman ay dahil sa malaking ibabaw ng dahon nito. Ang isang halaman ng mais ay kumonsumo ng 200 litro ng tubig sa panahon ng lumalagong panahon, at tubo - dalawang beses ang dami. Ang mga halaman ay pinipilit na bumuo ng isang malaking ibabaw ng dahon upang makatanggap kinakailangang halaga CO 2 para sa photosynthesis, sa kabila ng mababang nilalaman nito (0.045%) sa atmospera.
Halimbawa, ang isang 8 taong gulang na puno ng mansanas ay maaaring mag-assimilate ng hanggang 50 g ng CO 2 bawat araw na may intensive photosynthesis. Ito ay sumisipsip ng ganitong halaga ng CO 2 mula sa 300,000 litro ng hangin.
Ang pangangailangan na ubusin ang tubig sa maraming dami ay dahil din sa ang katunayan na ang pagsipsip ng sikat ng araw ng mga halaman ay dapat na humantong sa isang pagtaas sa temperatura, na maaaring maging sanhi ng coagulation ng mga protina. Sa pamamagitan ng pagsingaw ng tubig, pinababa ng halaman ang temperatura ng katawan nito.
Para sa karamihan ng mga halaman, ang pagkatuyo ay nakamamatay, kaya ang pagkonsumo ng tubig ay dapat na maiugnay sa kita. Ang mga lumot at lichen lamang ang makatiis ng mahabang kawalan ng tubig at makatiis sa oras na ito sa isang estado ng pagkatuyo.
Samakatuwid, sa sandaling lumipat ang mga halaman mula sa tubig papunta sa lupa, isang mahabang proseso ang nagsimulang bumuo ng mga adaptasyon upang mapanatili ang balanse ng tubig: una, upang mabilis na makapasok, at ikalawa, upang mabawasan ang basura ng mga terrestrial na organo; pangatlo, para sa cost-effective na transportasyon kasama ang stem.
Kapag sinusuri ang kaugnayan sa pagitan ng pag-agos at pag-agos ng tubig, tatlong mga kaso ang posible: ang pag-agos ay mas malaki kaysa sa pag-agos, katumbas o mas kaunti. Sa huling kaso, mayroong kakulangan sa tubig. Sa tanghali, ang kakulangan sa tubig ay maaaring umabot sa 5-10 at kahit na 25%. Isa sa mga kondisyon para sa normal na paggana ng mga halamang panlupa ay ang pagpapanatili ng mga kondisyon na walang mahaba at malalim na kakulangan sa tubig. Ito ay nangangailangan ng isang mahusay na binuo sistema ng ugat pagbibigay ng daloy ng tubig sa mataas na bilis.
Ang daloy ng tubig mula sa lupa hanggang sa mga ugat. Mas marami ang pag-agos ng tubig mula sa lupa mahirap na proseso kaysa sa simpleng pagsipsip ng tubig ng isang halaman na inilubog sa isang may tubig na solusyon ng mga ugat nito. Sa lupa mayroon tayong isang serye ng mga puwersa na sumasalungat sa pagsipsip na ito, na maaaring tinatawag na mga puwersang nagpapanatili ng tubig.
Ang lupa ay isang multiphase system na binubuo ng apat na pangunahing bahagi: solidong mga particle ng mineral, organikong bagay (humus), solusyon sa lupa at hangin sa lupa.
Ang mga particle ng mineral at humus ay bumubuo sa istraktura ng lupa, pinupuno ng tubig at hangin ang mga cavity ng istrakturang ito.
Ang kakayahan ng lupa na mapanatili ang tubig ay nakasalalay sa komposisyon at mga katangian nito. Ang isang tiyak na dami ng tubig ay bahagi ng mga mineral na bahagi ng lupa at hindi magagamit sa mga halaman (hygroscopic water). Ang iba't ibang mga clay mineral at heterogenous humic substance na mga colloid ay maaari ding magpanatili ng malaking halaga ng tubig ng hydration. Ang nasabing tubig ay may kondisyong tinatawag na nakatali (cellular) at mahirap ding ma-access. Ang tubig sa mga capillary ng lupa (capillary water) ay madaling hinihigop ng mga buhok ng ugat at mga cell sa ibabaw ng sucking zone ng ugat. Ang nasabing tubig ay itinuturing na libre.
Bilang karagdagan, sa lupa tayo ay nakikitungo sa isang solusyon, at hindi sa malinis na tubig; kaya ang solusyon mismo ay may lakas ng pagsuso. Kapag ang mga konsentrasyon sa lupa at sa mga sisidlan ng xylem ay nasa equilibrium, sa pangkalahatang kaso, ang pag-agos ng tubig ay dapat huminto.
Upang kalkulahin ang dami ng hindi magagamit na kahalumigmigan ng lupa, magpatuloy bilang mga sumusunod. Ang mga halaman ay pinatubo sa isang sisidlan na may mga dingding na hindi tinatablan ng tubig (salamin o metal) at pagkatapos na umunlad nang mabuti ang mga halaman, hindi na dinidiligan ang lupa at iniiwan sa isang madilim na lugar hanggang sa malanta ang halaman. Ang pagkalanta ay nagpapahiwatig na ang suplay ng tubig sa mga ugat ay huminto. Ang dami ng tubig na nananatili sa lupa sa oras ng pagkalanta ay hindi mapupuntahan ng halaman (dead stock). Ang dami ng hindi magagamit na tubig ay tinatawag na wilting factor o wilting moisture. Samakatuwid, ang wilting coefficient ay nagpapakilala sa kahalumigmigan kung saan nagsisimula pa lamang ang matatag na pagkalanta.
Sa puntong ito, mayroon pa ring tiyak na dami ng tubig sa lupa, bagaman maliit, ngunit sa ilang lawak ay magagamit para sa halaman. Sa ilalim ng mga kondisyong ito, kahit na ang isang napakalanta na halaman ay nabubuhay kung sinimulan mo itong diligin. Ang kumpletong pagkamatay ay nangyayari lamang kapag ang halumigmig ay mas mababa kaysa sa wilting coefficient.
Kaya, ang wilting moisture ay ang mas mababang limitasyon ng moisture range kung saan posible ang paglago ng halaman.
Kapag ang tubig ay pumasok sa tuyong lupa, ito ay nasisipsip nang napakabilis. Pagkatapos, ang rate ng pagpasok ng tubig sa mas mababang mga horizon ay nagiging mabagal. Kapag ang rate ng pababang daloy ng tubig ay bumaba nang husto, ang kahalumigmigan ng lupa ay umabot sa isang antas na tinatawag na field capacity.
Ang kahalumigmigan ng lupa na magagamit ng halaman ay tumutukoy sa dami ng tubig na naipon sa lupa mula sa antas ng kahalumigmigan ng matatag na pagkalanta hanggang sa kapasidad ng patlang.
Radial na transportasyon ng tubig. Upang magpatuloy sa isang pag-uusap tungkol sa radial na transportasyon ng tubig, kailangan mong maikling alalahanin karaniwang mga tampok mga istruktura ng ugat, (bagaman sa detalye ang istraktura ng mga ugat sa iba't ibang uri iba-iba ang mga halaman).
Ang parehong uri ng halaman ay may gitnang silindro (stele), na naglalaman ng mga vascular bundle at (stelar) parenkayma. Sa tulong ng mga vascular bundle (phloem), ang isang koneksyon ay pinananatili sa pagitan iba't ibang parte halaman at mga site ng carbohydrate synthesis. Ang mga vascular bundle (xylem) ay nagdadala ng tubig at mga dissolved substance mula sa ugat hanggang sa shoot. Kung lumipat ka sa paligid, pagkatapos ay mayroong isang recycle - isang solong hilera na layer ng mga cell. Pagkatapos ay darating ang endoderm, isa ring layer ng mga cell, lalo na mahusay na inangkop upang magsilbi bilang isang hadlang na naghihiwalay sa mga conductive tissue mula sa cortex. Matapos matapos ang pagpapahaba ng cell, ang isang layer ng materyal ay ipinahayag sa radial at transverse shell ng endoderm, na nagbibigay ng positibong reaksyon sa lignin. Ito ang tinatawag na Casparian belt. Mahigpit na nililimitahan ng mga banda ng Casparian ang paggalaw ng tubig at mga ion sa mga lamad ng cell. Kaya, epektibo nilang hinaharangan ang apoplastic transport.
Karamihan sa mga ugat ay inookupahan ng mga selula ng bark. Sila (kasama ang epidermis) ay bumubuo ng 86–90% ng lugar. Ang mga cell ng cortex ay pinahabang parallel sa pangunahing axis; isang manipis na layer ng cytoplasm (1-5 μm) ang pumapalibot sa gitnang vacuole sa kanila, na sumasakop sa ~ 90% ng dami ng protoplast. Ang mga pangalawang pagbabago sa mga lamad ng cell ng cortex ay nabawasan sa pagtitiwalag ng selulusa; lamang sa ilang mga species lignification nangyayari. Sa pangkalahatan, ang mga shell ay nagpapanatili ng permeability. Mayroon silang maraming mga pores na maaaring punan, depende sa mga kondisyon, na may hangin o tubig.
Sa paligid ng ugat ay ang epidermis (rhizodermis). Ito ang panlabas na tisyu ng ugat, na binubuo ng mga pinahabang siksik na mga selula. Ang mga lamad ng mga selulang ito ay maaaring sumailalim sa mga pagbabagong nauugnay sa pagtitiwalag ng cutin, isang sangkap na lumalaban sa tubig.
Ang mga panlabas na tangential lamad ng mga selula ng rhizodermis ay nakakapag-unat nang malakas at bumubuo ng tubular outgrowths, ang tinatawag na root hairs. Tandaan na ang mga cell na maaaring bumuo ng mga ugat na buhok ay tinatawag na trichoblast, at ang mga hindi kayang bumuo ay tinatawag na atrichoblast.
Kaya, ang tubig na pumasok sa ugat ng buhok o ibang root cell, sa tulong ng isa sa mga mekanismo na aming tinalakay, ay gumagalaw sa xylem vessels, ibig sabihin, mayroong isang radial na paggalaw ng tubig sa pamamagitan ng mga tisyu ng ugat.
Ang paggalaw ng tubig sa isang ugat na buhok o ibang cell ay nagsisimula sa pakikipag-ugnayan sa cell lamad, pagkatapos ang tubig ay maaaring dumaan sa plasmalemma at madala mula sa protoplast ng isang cell patungo sa protoplast ng isa pa sa pamamagitan ng plasmodesmata. Sa kasong ito, ang tubig ay gumagalaw kasama ang symplast. Gayunpaman, ang tubig ay hindi makapasok sa symplast, ngunit nananatili sa lamad ng cell at gumagalaw kasama ang mga ito sa pamamagitan ng mga tisyu ng ugat hanggang sa gitnang silindro. Ito ang apoplastic pathway. Ngunit sa kahabaan ng apoplast, ang tubig ay maaaring lumipat lamang hanggang sa endoderm (hanggang sa mga banda ng Casparian). Samakatuwid, ang landas ng tubig kasama ang apoplast ay humihinto dito. Para sa karagdagang paggalaw, dapat itong pumasok sa symplast.
Si D. Priestley ang unang nagpahayag ng barrier function ng endoderm noong 1920. Kamakailan lamang, ang function na ito ay paulit-ulit na nakumpirma.
Ang pangangailangan na ilipat ang paggalaw ng tubig mula sa apoplastic patungo sa symplastic na landas ay napakahalaga, dahil ang proseso ng paggalaw sa kahabaan ng buhay na protoplasm ay maaaring kontrolin ng cell mismo, sa kaibahan sa paggalaw ng tubig sa mga dingding ng cell.
Para makapasok ang tubig sa ugat, kinakailangan na ang potensyal ng tubig ng rhizodermis ay mas mababa kaysa sa potensyal ng tubig ng solusyon sa lupa; sa kasong ito, ang tubig ay magsisimulang dumaloy sa mga selula.
Ang lupa ay maaaring ituring bilang isang reservoir, ang dami ng tubig kung saan ito ay tumataas o bumababa. Kung, pagkatapos ng ulan, ang lupa ay nasa isang estado ng kapasidad sa bukid, kung gayon ang potensyal ng tubig nito ay malapit sa zero; madaling pumasok ang tubig sa mga ugat. Habang natutuyo ang lupa, bumababa ang potensyal ng tubig nito.
Ang potensyal ng tubig ay pinakamataas sa lupa, medyo mas mababa sa mga selula ng ugat, at pinakamababa sa mga selula na katabi ng epidermis ng dahon.
Ang unang dahilan para sa paglitaw ng isang potensyal na tubig na gradient ay ang aktibong pagpasok ng mga asin at ang kanilang aktibong paggalaw mula sa isang cell patungo sa isa pa. Ang pangalawa ay ang mga buhay na selula ng parenkayma ng gitnang silindro ay naglalabas ng natutunaw na mga organikong at mineral na sangkap sa mga sisidlan at sa gayon ay nagpapanatili ng potensyal na tubig na gradient sa mga sisidlan. Pagpasok sa mga xylem vessel, bumubuo sila ng osmotic gradient dito.
Ang potensyal na kemikal ng tubig kung saan matatagpuan ang mga ion na ito ay mas mababa kaysa sa potensyal ng tubig sa isang dilute na solusyon sa asin. panlabas na kapaligiran. Samakatuwid, ang tubig ay dapat lumipat kasama ang potensyal na kemikal na gradient patungo sa root xylem.
Ngayon, ang isang hypothesis ay iniharap, na may isang pang-eksperimentong batayan, na ang mga ugat ay may isang dalubhasang mekanismo para sa aktibong pumping ng tubig (water pump), ang pagpapatakbo nito ay hindi nakasalalay sa pag-agos ng mga ions.
Sa katunayan, kailangan ang enerhiya para makapasok ang tubig sa ugat; samakatuwid, ang prosesong ito ay nakasalalay sa bilis ng aerobic respiration ng mga ugat, ang pangunahing pinagmumulan ng ATP. Kaya, ang isa sa mga pangunahing salik na tumutukoy sa daloy ng tubig ay O 2 . Samakatuwid, ang mga kondisyon na may kakayahang sugpuin ang paghinga ay mabilis na nakakabawas sa daloy ng tubig. Bilang halimbawa, isaalang-alang ang sumusunod na kababalaghan: umulan nang malakas, maraming tubig ang naipon sa mababang lugar, at ang halaman ay nalalanta. Bakit? Ang labis na tubig sa lupa ay pinilit ang hangin mula dito, ang supply ng oxygen sa ugat ay mahirap, ang paghinga ay pinigilan. Kaagad mayroong pagbagal sa daloy ng tubig sa root system. Ipinapaliwanag ng halimbawang ito kung bakit hindi maganda ang pag-unlad ng mga halaman at namamatay pa sa mga lupang binaha ng tubig.
Ang pagbaha ay humahantong hindi lamang sa pagbaba ng halaga ng O 2 , kundi pati na rin sa pagtaas ng konsentrasyon ng CO 2 sa lupa, na pumipinsala sa mga lamad ng buhok ng ugat; mayroong isang pagbawas sa daloy ng tubig, na kung saan ay nakumpirma sa pamamagitan ng pagsugpo sa pagpapalabas ng katas.
Ang daloy ng tubig sa ugat ay depende rin sa temperatura ng lupa. Halimbawa, sa malamig na marshy soils, sa kabila ng malaking dami ng tubig, ang mga halaman ay nakakaranas ng kakulangan ng tubig, dahil sa mababang temperatura ang paghinga ay pinipigilan at ang supply ng enerhiya sa mga ugat ay nagambala. Sa ilalim ng mga kondisyong ito, ang mga halaman ay nagsisimulang bumuo ng isang xeromorphic na istraktura (maliit na mga selula, maraming stomata, atbp.), Na katangian ng mga tuyong lugar.
Ang estado ng mga halaman kung saan hindi makapasok ang tubig, sa kabila ng malaking halaga nito sa kapaligiran, ay tinatawag pisyolohikal na tagtuyot.
Ang iba't ibang mga pollutant ay maaari ding makaapekto sa paggamit ng tubig, lalo na sa pamamagitan ng pagpigil sa paghinga.
Ang pangangailangan na sumipsip ng isang malaking halaga ng tubig kahit na sa ilalim ng mga kondisyon ng limitasyon nito, halimbawa, sa mga kondisyon ng tagtuyot, ay humahantong sa katotohanan na ang halaman ay bumubuo ng isang malaking sistema ng ugat. Bilang resulta, ang mga ugat ay tumagos sa lupa nang mas malalim. Sa trigo, ang haba ng ugat ay umabot sa 90 cm, sa alfalfa - 120 cm Kung ipagpalagay natin na ang lalim ng arable horizon ay 20-25 cm, kung gayon ang karamihan sa root system ay matatagpuan sa ibaba. Ang laki ng root system ay nailalarawan hindi lamang sa lalim ng kanilang pagtagos sa lupa. Ang pangkalahatang ibabaw ay may malaking kahalagahan.
Ang tubig sa lupa ay gumagalaw nang napakabagal: sa buwan na ito ay nagkakalat ng hindi hihigit sa 30 cm. Ang paggalaw ng dulo ng ugat sa lupa ay nauuna sa paggalaw ng tubig. Kaya, hindi ang tubig ang gumagalaw patungo sa ugat, ngunit ang ugat patungo sa tubig sa proseso ng paglaki. Ang paglaki ay ang una at pinakamahalagang katangian ng mga ugat bilang isang organ na sumisipsip ng tubig.
Sa tuyo na mga kondisyon, ang isang root system ay nabuo 3-4 beses na mas malaki kaysa sa mga basa.
Sumasanga at mabilis na paglaki tulungan ang ugat na lumipat patungo sa tubig, ngunit, sa kabilang banda, tubig - kinakailangang kondisyon paglago. Tulad ng nakikita mo, hindi sa cellular, ngunit sa antas ng organismo, nahaharap tayo sa isang halimbawa ng feedback, na sumasailalim sa mga proseso ng regulasyon.
Bilang isang organ na sumisipsip ng tubig, ang ugat ay may isa pang mahalagang pag-aari - positibong hydrotropism, na nangangahulugan na sa kakulangan ng tubig, ang mga lumalagong bahagi ng mga ugat ay yumuko patungo sa mas mahalumigmig na mga lugar ng lupa.
Kaya, ang root system ay isang dalubhasang organ para sa paggamit ng tubig.
Ang paglago ng ugat ay kadalasang higit sa paglaki ng mga organo sa lupa. Ito ay lubhang mahalagang katangian nauugnay sa katotohanan na ang ugat ay dapat magbigay ng mga pangangailangan ng tubig ng bumubuo ng halaman.
Gayunpaman, ang isang cell sa anumang organ na hindi puspos ng tubig ay maaari ding sumipsip ng tubig sa sandaling ito ay madikit dito. Samakatuwid, ang mga dahon, lalo na ang mga lanta, kapag inilubog sa tubig, ay hinihigop ito nang masigasig; sa kabilang banda, sa kabila ng cuticle, ang tubig ay maaaring pumasok sa ibabaw ng dahon. Tanging ang tuyong kutikyol ay naipakita na halos hindi tinatablan ng tubig; kapag basa, ito ay namamaga at nagiging permeable, kaya ang mga dahon na nabasa ng ulan o hamog ay maaaring sumipsip ng hanggang 25% ng tubig na nahuhulog sa kanila. Mayroon itong praktikal na halaga kapag nagdidilig ng mga halaman sa pamamagitan ng pagwiwisik.
Long distance transport ng tubig. Sa pagsasalita tungkol sa paggalaw ng tubig sa pamamagitan ng halaman, ang transportasyon sa mga tisyu ng isang organ, na tinatawag na malapit (radial), at transportasyon sa pagitan ng mga indibidwal na organo, na tinatawag na malayo, ay nakikilala. Mayroong isang makabuluhang pagkakaiba sa pagitan nila. Ang short-range na transportasyon ay dumaan sa mga hindi espesyal na tissue, at para sa long-range na transportasyon, ang mga halaman ay may mga espesyal na conductive tissue. Kaya, ang landas na dinaraanan ng tubig mula sa ugat na buhok patungo sa evaporating na selula ng dahon ay nahahati sa dalawang bahagi: iba sa haba, istraktura, at mga katangiang pisyolohikal. Ang unang bahagi ay binubuo ng mga buhay na selula at maliit ang sukat (milimetro o mga fraction ng isang milimetro). Ito ay dalawang maikling seksyon - isa - sa ugat, mula sa ibabaw nito na may mga ugat na buhok hanggang sa mga sisidlan na nasa gitnang silindro nito; ang pangalawa - sa dahon, mula sa mga sisidlan na bahagi ng conductive bundle, at chlorenchyma evaporating tubig sa intercellular space. Ang ikalawang bahagi ng landas ay mga sisidlan, mga tracheid, na mga patay na tubo. Sa mala-damo na mga halaman, ang kanilang haba ay ilang sentimetro, habang sa mga puno umabot ito ng ilang metro at kahit sampu-sampung metro.
Ang mga elemento ng tubig at mineral ay inihahatid sa bawat cell ng aerial na bahagi ng halaman dahil sa pataas na agos sa pamamagitan ng xylem. Mayroon ding pababang daloy ng phloem ng mga solusyon mula sa mga dahon hanggang sa mga ugat. Ang pababang phloem current ay nabuo sa mga cell ng leaf mesophyll, kung saan ang bahagi ng tubig na kasama ng xylem current ay dumadaan mula sa mesophyll cell membranes patungo sa phloem endings.
Ang tubig mula sa mga selula ng dahon at direkta mula sa mga sisidlan ng xylem ay pumapasok sa phloem kasama ang osmotic gradient na nangyayari dahil sa mga asukal at iba pang mga organikong compound na naipon sa mga selula ng phloem, na nabuo sa panahon ng photosynthesis.
Ang pababang daloy ng phloem ay naghahatid ng mga organikong sangkap sa mga tisyu ng ugat, kung saan ginagamit ang mga ito sa metabolismo. Sa mga ugat, ang mga dulo ng pagsasagawa ng mga bundle ng mga elemento ng phloem, tulad ng sa dahon, ay matatagpuan malapit sa mga elemento ng xylem, at ang tubig ay muling pumapasok sa xylem kasama ang osmotic gradient at gumagalaw paitaas. Kaya, mayroong isang palitan ng tubig sa pagsasagawa ng sistema ng mga ugat at dahon (na parang isang cycle).
Ang daloy ng tubig sa pamamagitan ng mga sisidlan ng xylem ay humahantong sa katotohanan na kapag ang tangkay ng isang halaman ay pinutol sa isang maikling distansya mula sa lupa, pagkatapos ng ilang oras, ang katas ay nagsisimulang tumayo mula sa dulo ng mga sisidlan, na tinatawag na apiary. Ang phenomenon na ito ay tinatawag na "weeping plants."
Ang puwersa na nag-aangat ng katas sa mga sisidlan ay tinatawag na presyon ng ugat. Maaaring masukat ang presyon ng ugat kung ang isang tubo na inilagay sa isang putol na tangkay ay konektado sa isang pressure gauge. Ang halaga ng presyon ng ugat ay hindi pare-pareho. Sa ilalim ng pinakamainam na kondisyon, ito ay 2-3 bar. Sa ilalim ng ilang partikular na kundisyon, ang isang equilibrium ay naabot sa pagitan ng dami ng sap na ginawa at ng dami ng tubig na ibinibigay, kaya ang presyon ng ugat, o ang dami ng sap na ginawa, ay maaaring sumasalamin sa kapasidad ng pagsipsip ng mga ugat. Kaya, ang mga aktibong makina ng paunang pagtaas ng kasalukuyang tubig (presyon ng ugat) ay mga nabubuhay na selula na katabi ng mas mababang dulo ng sistema ng pagsasagawa ng mga halaman - ito ang mga selula ng root parenchyma - ang mas mababang terminal ng makina ng daloy ng tubig.
Ang mekanismo ng presyon ng ugat ay pinaniniwalaan na nakabatay sa pagkilos ng mga contractile na protina, ang pagpapaandar nito ay pinaniniwalaang ginagawa ng microfibrils ng P-proteins.
Sa ilang lawak, ang guttation ay maaaring magsilbing ebidensya ng aktibong pagpapalabas ng tubig.
Gayunpaman, kung ang halaman ay hindi patuloy na nawawalan ng tubig bilang resulta ng transpiration, kung gayon ang mga selula ng buhok ng ugat ay mabilis na mabubusog ng tubig, at ang supply nito ay titigil. Samakatuwid, ang isa sa mga dahilan para sa paglitaw ng isang potensyal na gradient ng tubig ay ang pagsingaw ng tubig ng mga organo sa itaas ng lupa.
Kung mas matindi ang pagsingaw ng tubig ng mga selula ng dahon, mas mabilis itong papasok sa mga selula ng ugat at mas mabilis na madala sa halaman. Ang pagkawala ng mga molekula ng tubig sa tuktok ng haligi ng tubig sa pamamagitan ng pagsingaw ay nagiging sanhi ng pag-agos ng tubig pataas sa pamamagitan ng mga sisidlan ng xylem upang maalis ang pagkawala. Ang paggalaw ng tubig na ito na dulot ng transpiration ay tinatawag na transpiration current. Tinutukoy nito ang daloy ng tubig mula sa lupa patungo sa halaman kasama ang parehong water potential gradient. Dahil sa transpiration, ang potensyal ng tubig sa tuktok ng halaman ay mas mababa kaysa sa base.
Ang mga aktibong makina ng daloy ng tubig dahil sa transpiration ay mga buhay na selula na katabi ng itaas na dulo ng buong sistema ng pagsasagawa ng halaman - ang mga selula ng parenkayma ng dahon. Tinatawag silang top end motor ng tubig na kasalukuyang.
Ang mekanismo ng pagpapatakbo ng upper end motor ay simple at batay sa mga sumusunod. Ang kapaligiran ay karaniwang undersaturated na may singaw ng tubig at samakatuwid ay may negatibong potensyal na tubig. Sa isang relatibong halumigmig na 90%, ito ay 140 bar. Sa karamihan ng mga halaman, ang potensyal ng tubig ng mga dahon ay mula 1 hanggang 30 bar.
Dahil sa malaking pagkakaiba sa mga potensyal ng tubig, nangyayari ang transpiration. Ang pagbawas sa dami ng tubig sa parenchymal cell ng dahon ay nagdudulot ng pagbawas sa aktibidad ng tubig sa loob nito at pagbaba sa potensyal ng tubig.
Ang kakulangan ng tubig ay unti-unting umabot sa mga ugat, at ang aktibidad ng tubig sa mga ito ay bumababa. Sa kasong ito, ang tubig ay nagmumula sa lupa hanggang sa ugat. Kaya, maaari itong tapusin na ang paggalaw ng tubig sa pamamagitan ng halaman, pati na rin ang pagpasok nito sa ugat, ay pangunahing nauugnay sa gradient ng potensyal ng tubig sa sistema ng lupa-halaman-hangin. Ang gradient na ito ay magiging mas malaki, mas maraming tubig ang mawawala sa mga selula ng dahon, ibig sabihin, mas malakas ang transpiration.
Magkaiba ang paggana ng dalawang makina. Sa karaniwan, ang upper end na motor ay nagkakaroon ng puwersa na 10-15 bar at higit pa, at ang mas mababang isa ay 2-3 bar. Ipinapakita nito na ang pangunahing papel sa pagpapalitan ng tubig ay kabilang sa itaas na makina. Gayunpaman, sa kawalan ng mga dahon sa mga puno sa taglamig at sa unang bahagi ng tagsibol, o pagkatapos ng tuyong panahon nangungunang papel sa paggalaw ng tubig ay gumaganap ang mas mababang engine. Ang lower terminal motor ay dapat gumanap ng isang malaking papel sa pagtaas ng tubig sa pamamagitan ng halaman sa ilalim ng mga kondisyon ng mas mataas na air humidity, kapag ang transpiration ay minimal.
Para sa upper end engine, ang pinagmumulan ng enerhiya ay ang araw, na nangangahulugan na ang nagniningning na enerhiya na hinihigop ng dahon ay ginagamit para sa pagsingaw.
Para sa lower terminal motor, ang pinagmumulan ng enerhiya ay paghinga. Ang enerhiya ng mga molekula ng ATP, na na-synthesize sa panahon ng paghinga ng mga selula ng ugat, ay ginugugol sa transportasyon ng mga ions sa cell, ibig sabihin, sa paglikha ng isang potensyal na gradient ng tubig. Ang regulasyong papel ng presyon ng ugat sa pagpapalitan ng tubig ng halaman ay eskematiko na ipinapakita sa fig. 4.12.
Kaya, ang upper end engine ay isang awtomatikong operating mechanism na sumisipsip ng tubig nang mas malakas, mas mabilis itong kumonsumo.
Ang pagpapatakbo ng upper at lower end motors ay madaling maipaliwanag ang pagtaas ng tubig ng ilang sampu-sampung sentimetro, kahit na metro. At paano ipaliwanag ang pagtaas ng tubig sa sampu-sampung metro, at ang sequoia ay umabot sa taas na 140 metro? Ang mga sisidlan kung saan ang tubig ay gumagalaw para sa karamihan ng landas nito ay mga patay na tubo. Hindi sila makabuo ng lakas para magbuhat ng tubig. Ang teorya ng adhesion, na iminungkahi ng English researcher na si G. Dixon noong 1921, ay nakakatulong upang masagot ang tanong na ito. Alinsunod sa teoryang ito, ang mga tuluy-tuloy na mga thread ay nabuo sa mga sisidlan, na dumadaan mula sa mga cell ng root parenchyma hanggang sa mga cell ng parenchyma ng dahon. Ang puwersa na nagiging sanhi ng mga molekula ng tubig na sumunod sa isa't isa ay tinatawag na cohesive force (cohesion). Ang tuluy-tuloy na mga hibla ng tubig ay nabuo sa pamamagitan ng mga bono ng hydrogen. Gayunpaman, ang mga filament ng tubig ay nakakabit din sa mga pader ng sisidlan (adhesion) na may puwersang 300–350 bar. Ang lahat ng ito ay nagpapahintulot sa mas mababang at itaas na dulo ng mga motor na itaas ang tubig kasama ang baras sa taas na 140 m.
Matapos ang pagdating ng teoryang ito, paulit-ulit na iginuhit ng mga anatomist ang atensyon ng mga mananaliksik sa pagbuo ng mga bula ng hangin, na dapat masira ang pagdirikit sa pagitan ng mga molekula ng tubig sa mga sisidlan. Gayunpaman, sa kaso ng isang pansamantalang pagbubukod ng isang sisidlan, ang tubig ay gumagalaw kasama ang mga siding (iba pang mga sisidlan) o ang apoplast, at ang mga bula ng hangin ay unti-unting nalulutas sa partisipasyon ng mga buhay na selula.
Ang paggalaw ng tubig mula sa ugat hanggang sa dahon kasama ang mga patay na sisidlan, na nag-aalok ng minimal na pagtutol sa agos ng tubig, ay isa sa mga natuklasan ng kalikasan, na binubuo ng mga sumusunod. Ang mga selula ng mga sisidlan at tracheid ay pinahaba, wala silang buhay na nilalaman, sila ay walang laman sa loob, i.e. sila ay mga simpleng tubo. Ang lignified pangalawang cell lamad ay sapat na malakas upang mapunit, na makatiis sa malaking pagkakaiba sa presyon na nangyayari kapag ang tubig ay tumaas sa tuktok ng malalaking puno. Ang dulo, at kung minsan ang mga dingding sa gilid ng mga segment ng mga sisidlan, ay butas-butas; Ang mga sisidlan, na binubuo ng mga segment na pinagsama sa mga dulo, ay bumubuo ng mga mahahabang tubo kung saan ang tubig na may mga mineral ay madaling dumaan. Walang mga butas sa mga tracheid, at ang tubig, upang makarating mula sa isang tracheid patungo sa isa pa, ay dapat dumaan sa kanilang mga dulong dingding; ngunit ang mga tracheid ay napakahabang mga selula, at samakatuwid ang istrukturang ito ay napakahusay ding iniangkop para sa pagdadala ng tubig.
Sa proseso ng ebolusyon ng halaman, ang paglitaw ng mga halaman sa lupa, ang korona na kung saan ay matatagpuan medyo malayo sa lupa, ay naging posible dahil sa pagbuo ng isang mataas na dalubhasang sistema ng pagsasagawa. Ang kahalagahan ng pagbagay na ito ay binibigyang diin din ng pangalan ng mga halaman mismo - vascular.
Bilang karagdagan sa pagkakaiba sa mga mekanismo ng pagkilos, mayroong kumpletong pagkakapare-pareho sa pagpapatakbo ng dalawang dulo ng motor. Tumutugon ang dahon sa pagkilos ng anumang kadahilanan sa kapaligiran na maaaring sugpuin ang pagpapatakbo ng mas mababang motor sa pamamagitan ng pag-activate ng transpiration, at kabaliktaran. Ito ay isang biologically napakahalagang pagbagay, kahit na sa unang sulyap ito ay mukhang kabalintunaan: ang suplay ng tubig ay lumalala, at ang dahon ay tumutugon sa hindi kanais-nais na kadahilanan na ito hindi sa pamamagitan ng pagsugpo, ngunit, sa kabaligtaran, sa pamamagitan ng pagtaas ng transpiration. Ang pagtaas ng transpiration sa kasong ito ay inilaan upang pasiglahin ang daloy ng tubig sa ugat.
Ngayon ay may isang view na ang halaman ay may isang espesyal na sistema ng regulasyon - hydrodynamic. Sa ilalim ng kontrol nito ay ang rehimen ng tubig, ang pagpapanatili ng homeostasis ng tubig, pati na rin ang ilang iba pang mga function, sa partikular, photosynthesis. Napakasensitibo ng hydrodynamic regulatory system. Ito ay kumikilos na may napakakaunting pagkawala ng tubig mula sa dahon (0.06% ng orihinal na halaga) at pinipigilan ang karagdagang pag-aalis ng tubig.
Ang paghahatid ng signal sa dahon ay nangyayari sa pamamagitan ng tuluy-tuloy na daloy ng tubig, at ang pang-unawa ay isinasagawa ng stomata at photosynthetic apparatus. Ang receptor para sa mga pagbabago sa mga kondisyon ng lupa, na pumipigil sa pag-agos ng tubig, ay malamang na ang mga lamad ng root endodermal cells.
Ang hydrodynamic regulatory system ay nagbibigay-daan sa planta na tumugon nang napakabilis sa mga panlabas na pagbabago na posibleng hindi kanais-nais para sa homeostasis ng tubig.
Ang mga halaman sa lupa ay nahaharap sa isang mahirap na problema: sa isang banda, dapat silang magkaroon ng isang sapat na binuo na ibabaw upang epektibong sumipsip ng sikat ng araw at CO 2 , at sa kabilang banda, habang tumataas ang ibabaw, tumataas ang pagkawala ng tubig. Nalutas ng mga halaman ang problemang ito iba't ibang paraan: una, ang suplay ng tubig ay tumataas dahil sa paglaki ng mga ugat at pag-unlad ng hypertrophied absorption surface. Pangalawa, ang pagkawala ng tubig ay nagiging mas mabagal dahil sa ang katunayan na ang mga mesophilic na selula ay pinaghihiwalay mula sa kapaligiran sa pamamagitan ng isang cuticle na naglalaman ng wax. Pangatlo, ang kontradiksyon sa pagitan ng pangangailangang sumipsip malaking dami CO 2 at sa parehong oras bawasan ang halaga ng evaporated tubig, halaman magpasya gamit ang isang oscillatory mekanismo.
Mga paraan ng malapit at malayong transportasyon, ang mekanismo ng paggalaw ng tubig sa pamamagitan ng halaman (potensyal na gradient ng tubig, mga puwersang nagtutulak ng pataas na daloy ng tubig sa halaman, upper at lower end motors, adhesion at cohesion na proseso).
Gitnang transportasyon- ito ang paggalaw ng mga ion, metabolite at tubig sa pagitan ng mga selula at tisyu (kumpara sa transportasyon ng lamad sa bawat selula). malayuang transportasyon- ang paggalaw ng mga sangkap sa pagitan ng mga organo sa buong halaman.
Ang transportasyon ng mga sangkap sa isang halaman ay isinasagawa sa pamamagitan ng anumang mga tisyu at sa pamamagitan ng conductive bundle na dalubhasa para sa layuning ito. Kaugnay nito, ang paggalaw ng tubig at mga solute sa pamamagitan ng anumang mga tisyu ay maaaring mangyari: a) kasama ang mga dingding ng cell, ibig sabihin, kasama ang apoplast, b) kasama ang cytoplasm ng mga cell na konektado sa bawat isa sa pamamagitan ng plasmodesmata, ibig sabihin, kasama ang symplast, c) posibleng kasama ang endoplasmic reticulum na may partisipasyon ng plasmodesmata.
Ang paggalaw ng tubig at mga sangkap kasama ang conductive bundle ay kinabibilangan ng transportasyon kasama ang xylem ("pataas na kasalukuyang" - mula sa mga ugat hanggang sa mga organo ng shoot) at kasama ang phloem ("pababang kasalukuyang" - mula sa mga dahon hanggang sa mga zone ng pagkonsumo ng nutrisyon o ang kanilang deposisyon sa stock). Ang mga metabolite ay dinadala din sa kahabaan ng phloem sa panahon ng pagpapakilos ng mga reserbang sangkap.
radial na transportasyon. Sa pamamagitan ng pagsasabog at metabolic proseso Ang mga ion ay pumapasok sa mga cell wall ng rhizodermis at pagkatapos ay lumipat sa core parenchyma patungo sa conductive bundle. . Ang paggalaw na ito ay nangyayari kapwa sa kahabaan ng mga pader ng cell - ang apoplast, at kasama ang symplast.
Ang paggalaw ng ion sa pamamagitan ng apoplast nangyayari dahil sa diffusion at exchange adsorption kasama ang gradient ng konsentrasyon at pinabilis ng daloy ng tubig. Paggalaw ng mga mineral sa pamamagitan ng symplast Isinasagawa ito dahil sa paggalaw ng cytoplasm, at sa pagitan ng mga cell - kasama ang plasmodesmata. Ang direktang paggalaw sa kahabaan ng symplast ay pinadali ng mga gradient ng konsentrasyon ng mga sangkap. Ang mga gradient na ito ay lumitaw dahil sa ang katunayan na ang mga sangkap na pumasok sa cell ay kasama sa mga proseso ng metabolic at bumababa ang kanilang konsentrasyon.
Ang pagsasabog ng mga ion at molekula sa pamamagitan ng maliwanag na libreng espasyo ng mga selula ay nagambala sa antas ng endoderm. Ang tanging paraan para sa karagdagang paggalaw ng mga sangkap sa pamamagitan ng endoderm ay ang transportasyon kasama ang symplast, na sinisiguro ng metabolic control ng paggamit ng mga sangkap. Pagkakaroon sa endoderm pumasa sa mga cell, kung saan ang mga banda ng Casparian ay kulang sa pag-unlad o wala, ay nagbibigay-daan sa isang maliit na bahagi ng hinihigop na mga sangkap na makatakas sa metabolic control.
Symplastic transport ay ang pangunahing isa para sa maraming mga ions. Kasabay nito, ang mga compound na naglalaman ng nitrogen, carbon, phosphorus, at, sa mas mababang lawak, ang sulfur, calcium, at chlorine ay sumasailalim sa aktibong metabolismo. Ang ibang mga ion ay halos hindi napapailalim sa metabolic control.
Ang isang mahalagang papel sa symplastic na transportasyon ng mga sangkap ay nilalaro ni mga vacuoles. Sa isang tiyak na lawak, nakikipagkumpitensya sila sa mga sisidlan ng xylem para sa mga hinihigop na sangkap at sa gayon ay kumikilos bilang isang regulator ng daloy ng mga sangkap sa mga sisidlan. Ang prosesong ito ay nakasalalay sa antas ng saturation ng vacuolar juice na may mga dissolved substance. Sa isang pagbawas sa konsentrasyon ng mga sangkap sa cytoplasm, maaari silang muling umalis sa mga vacuoles, kaya kumakatawan sa isang reserbang pondo ng mga sustansya. Ang pagsipsip ng mga ion sa pamamagitan ng mga vacuole ay binabawasan ang kanilang konsentrasyon sa symplast at tinitiyak ang paglikha ng isang gradient ng konsentrasyon na kinakailangan para sa kanilang transportasyon sa pamamagitan ng symplast.
Paano pumapasok ang mga ion sa mga patay na sisidlan ng xylem, ibig sabihin, paano ito na-load?
xylem sap ay isang solusyon na pangunahing binubuo ng mga di-organikong sangkap. Gayunpaman, sa katas na dumadaloy mula sa hemp xylem kapag ang itaas na bahagi ng tangkay ay inalis, iba't ibang mga nitrogenous compound (amino acids, amides, alkaloids, atbp.), Mga organic na acid, organophosphorus esters, mga compound na naglalaman ng sulfur, isang tiyak na halaga ng asukal. at polyhydric alcohols, pati na rin ang phytohormones ay natagpuan din. Ang xylem sap ay maaari ding maglaman ng mas kumplikadong mga sangkap na nanggagaling dito mula sa mga vacuole at cytoplasm ng mga elemento ng tracheal na kumukumpleto sa kanilang pag-unlad.
Ang Xylem sap ay naiiba nang husto mula sa vacuolargo sa komposisyon. Halimbawa, ang nilalaman ng K + ion sa pea epicotyl vacuoles ay umabot sa 55-78 mmol/l, habang sa xylem sap ay 2-4 mmol/l lamang.
Ang paglo-load ng xylem ay nangyayari nang mas matindi sa zone ng mga ugat na buhok. Isa o dalawang bomba ang gumagana sa zone na ito. Ang pangunahing soybean ay naisalokal sa plasmalemma ng rhizodermal at bovine parenchyma cells. Ito ay dahil sa gawain ng H + -pumps, na mga H + -ATPases at proton-transferring redox chain. Sa bahaging ito ng ugat, ang mga cation at anion mula sa mga pader ng cell ay pumapasok sa cytoplasm. Ang tubig at mga mineral na asin ay dumadaan sa mga endoderm na selula na may mga bandang Casparian lamang sa kahabaan ng symplast. Sa mga selula ng parenchymal ng bundle, na direktang katabi ng mga tracheid o mga sisidlan, ang pangalawang bomba ay gumagana, na naglalabas ng mga mineral na pumapasok sa kanilang mga cavity sa pamamagitan ng mga pores sa mga dingding ng mga elemento ng tracheal. Salamat kay aktibong gawain Ang dalawang bomba sa mga tracheid at mga sisidlan ay nagpapataas ng osmotic na potensyal at, dahil dito, ang lakas ng pagsuso.
Kapag lumalaki ang mga halaman, ang pagtutubig ay walang alinlangan ang pinakamalaking hamon. Sa isang banda, ito ay isang simpleng operasyon. Ngunit upang matantya kung gaano karaming tubig ang kailangan para sa isang halaman, at upang matukoy ang oras ng pagtutubig, kinakailangan ang ilang karanasan. Kakaibang tila, ngunit bilang isang patakaran, ang labis na tubig ay dapat sisihin para sa pagkamatay ng mga halaman, at hindi nangangahulugang kakulangan nito. Lupa oversaturated sa tubig hinaharangan ang pag-access sa oxygen para sa mga ugat, at ang hindi gumagalaw na malamig na tubig ay isang kapaligiran na nakakatulong sa simula ng kanilang pagkabulok. Samakatuwid, sa karamihan ng mga kaso, anuman ang temperatura at panahon, ang pang-araw-araw na pag-spray ng mga halaman ay mas mainam kaysa sa pagtutubig ng lupa.
enerhiyang solar nagiging sanhi ng pagkilos ng mga espesyal na organo ng dahon - mga chloroplast. Salamat sa kanila, ang mga dahon ay gumagamit ng carbon dioxide upang synthesize ang mga karbohidrat, na sa turn, sa pamamagitan ng iba't ibang mga reaksiyong kemikal, ay nagiging " materyales sa pagtatayo na bumubuo ng mga halaman.
Ang isang malusog na halaman ay kumukuha ng tubig mula sa lupa. Kasama nito, ang mga mineral ay tumaas kasama ang mga ugat, dinadala kasama ang tangkay, at pagkatapos ay kasama ang mga sanga na naabot nila ang mga dahon. Mula doon, ang tubig ay nagdadala ng mga sangkap na na-synthesize sa mga dahon sa iba pang mga organo ng halaman, na kung saan ay nakakatulong sa pag-unlad ng mga dahon at mga ugat.
Sa mga proseso ng paghinga at marami pang iba mga reaksiyong kemikal na nagaganap sa halaman, ang tubig ay sinisipsip. Samakatuwid, sa panahon ng paglago, maraming tubig ang kailangan. Ang tubig ay nagdadala din ng mga sustansya sa mga bahagi ng halaman na nangangailangan nito.
Pag-iwas sa Root Rotting
Alisin ang halaman mula sa palayok at alisin ang anumang may sakit na mga ugat - sila ay nagiging kayumanggi at nagiging malambot. Maingat na bunutin ang mga ugat. Kung ang panlabas na takip ay madaling natanggal, kung gayon ang ugat ay nabubulok. Ang magagawa lamang upang maibalik ang halaman ay putulin ang mga patay na ugat.

Pagdidilig
Ang kakayahang hatulan ang pangangailangan para sa pagdidilig ng isang halaman ay isang kalidad na nanggagaling sa karanasan na nakuha mula sa pagmamasid sa paglaki ng isang halaman sa paglipas ng panahon. Ang operasyon mismo ay simple. Mayroong ilang mga patakaran na tutulong sa iyo na maisagawa ang operasyong ito.
Ang halaman ay nangangailangan ng maraming tubig sa panahon ng paglago.. Nangyayari ito, bilang panuntunan, sa mainit na tagsibol at mga araw ng tag-init, lalo na kapag ang mga unang buds ay sumabog at ang halaman ay namumulaklak. Ngunit sa taglagas at taglamig, kapag bumaba ang temperatura, at ang karamihan sa mga halaman ay nagpapahinga, sa kabaligtaran, kaunting tubig ang kailangan. Ang isa pang bagay ay kapag ang halaman ay nasa isang napakainit na silid. Pagkatapos ay kailangan nilang madalas na natubigan, tulad ng ginagawa sa tag-araw.
Maraming mga halaman ang nangangailangan ng pahinga sa taglamig kung saan kailangan nila ng ilang tubig. At may ilan, tulad ng cacti at matabang babae, na kailangan lamang hugasan. Sa tagsibol, sa panahon ng malakas na paglaki, ang iyong mga halaman ay nangangailangan ng masaganang pagtutubig. Ilubog ang sisidlan na may halaman hanggang sa labi sa isang malaking lalagyan ng tubig. temperatura ng silid at panatilihin ito hanggang pagkatapos. hanggang basa ang ibabaw. Pagkatapos ay ilabas ito at hayaang maubos ang tubig. Ang isang katulad na operasyon ay maaaring gawin sa mga ampelous na halaman na nasuspinde nang mataas at hindi namin palaging masuri kung ang earthen ball ay mahusay na moistened.
Ang mga halaman ay pinakamahusay na natubigan sa umaga. upang ang labis na kahalumigmigan ay maaaring sumingaw sa araw. Kapag ang pagtutubig sa gabi, ang halaman sa pangkalahatan ay nananatiling basa sa buong gabi, at ang pagbaba ng temperatura ay nag-aambag sa pagbuo ng mga mapanganib na fungi ng peste. Maaari rin itong humantong sa pagkabulok ng ugat.
Isang halaman na ganap na napuno ang palayok ng mga ugat nito kailangang madidilig nang mas madalas.
Dalas ng pagtutubig tinutukoy din ang uri ng lupa. Ang mga clay soil ay sumisipsip ng tubig nang paunti-unti, ngunit dahan-dahan ding natuyo; mabuhangin - napakabilis, ngunit agad na nagiging tuyo muli. mabuhangin na mga lupa- ay perpekto.
Mga halaman sa clay pot, kumpara sa mga halaman sa mga plastik na kaldero, ay nangangailangan ng dalawang beses na mas maraming tubig. Ang dahilan nito ay ang luwad ay sumisipsip ng kahalumigmigan sa pamamagitan ng mga pores nito, na sumingaw sa mga dingding ng palayok.
Anumang mga halaman na may mabalahibo o pubescent na dahon, tulad ng saintpaulia at synningia, hindi madidiligan mula sa itaas, dahil ang mga basang dahon ay maaaring mabulok o mapasailalim sa mga fungal disease. Una sa lahat, mag-ingat sa mga cyclamen, dahil ang tubig ay tumagos sa gitna ng tuber, na maaaring maging sanhi ng pagkabulok ng mga tangkay ng dahon at mga tangkay ng bulaklak. Ang mga kaldero ay maaari lamang ilubog hanggang sa labi sa maligamgam na tubig sa loob ng mga 5 minuto. Pagkatapos ay dapat mong hayaan silang matuyo nang lubusan.
Iba't ibang halaman mula sa pamilyang bromeliad nangangailangan ng sariwang tubig sa gitna ng palayok kahit isang beses sa isang linggo. Ngunit ang mga epiphytic na halaman, tulad ng Tillandsia (Tillandsia), na natural na tumutubo sa mga piraso ng kahoy, ay magpapasalamat sa araw-araw na pag-spray ng tubig sa tag-araw at linggu-linggo sa taglamig. Ngunit bukod dito, bigyang-pansin din ang oras ng pagtutubig. Kung ang halaman ay nasa maaraw na lugar, iwasang matubigan ang mga bulaklak at dahon.

Ibabaw na crust
Ito ay nangyayari na ang ibabaw ng lupa ay nagiging matigas, matigas. Sa kasong ito, ang tubig ay nananatili sa ibabaw at hindi tumagos sa mga ugat. Alisin ang crust mula sa lupa. Mag-ingat na hindi makapinsala sa mga ugat.

Tuyong lupa
Ang pit, kung hindi pinananatiling basa, ay naninirahan, at kapag nagdidilig, ang tubig ay ganap na umaagos palabas ng palayok. Sa kasong ito, isawsaw ang halaman, kasama ang palayok, sa isang lalagyan ng tubig sa temperatura ng silid. Panatilihin ito doon hanggang sa huminto ang paglabas ng mga bula ng hangin sa lupa.
Mga Pagsusuri sa kahalumigmigan
Kung inilagay mo ang halaman sa isang clay pot, itali ito ng cotton thread sa isang bamboo stick at bahagyang kumatok sa kaldero. Ang isang malakas na tunog ay nagpapahiwatig na ang luad at ang lupa ay tuyo, samakatuwid ang halaman ay nangangailangan ng pagtutubig. Ang isang mapurol na tunog kapag hinahampas ang palayok ay nagpapahiwatig na ang lupa ay sapat na basa.
Kumuha ng palayok (luwad man o plastik) na may halaman. Kung nakita mong mabigat, ang lupa ay mamasa-masa, kung magaan, ang lupa ay tuyo. Ito ay isang medyo maaasahang pagsubok, ngunit nangangailangan ito ng ilang karanasan upang bumuo ng isang pamantayan para sa pinakamainam na timbang ng halaman. At para sa malalaking halaman, ito ay karaniwang hindi naaangkop.
Bago ang pagtutubig, maaari mong suriin ang kalagayan ng lupa sa pamamagitan lamang ng pagpindot sa ibabaw nito gamit ang iyong mga daliri. Kung ang ibabaw ng lupa ay tuyo, kung gayon ang halaman ay kailangang natubigan; kung ang lupa ay basa pa, pagkatapos ay maaari ka pa ring maghintay na may pagtutubig.
Kung hindi ka nagtitiwala sa mga naturang pagsubok, bumili aparato sa pagsukat ng kahalumigmigan. Ang metal probe ng aparato ay nahuhulog sa lupa. Ang isang tagapagpahiwatig na nilagyan ng isang arrow ay naka-install sa itaas na bahagi nito; ito ay nagpapahiwatig ng kalagayan ng lupa: "basa", "basa", "tuyo". Sa pamamagitan nito, madaling malaman kung ang halaman ay kailangang diligan.
Anong tubig ang gagamitin sa irigasyon?
Ang ordinaryong tubig mula sa gripo ay tinatanggap para sa karamihan ng mga halaman, kahit na ito ay matigas, na may mataas na nilalaman ng dayap. Ngunit gayon pa man, para sa mga halaman tulad ng azalea, erica, striped echmea (Aechmea fasciata), gumamit ng tubig-ulan. Ang mga halaman na ito ay hindi pinahihintulutan nang mabuti ang dayap. Ngunit gayon pa man, gamit tubig sa gripo, dapat mong hayaan itong tumayo ng kaunti bago ang pagdidilig upang ito ay makakuha ng ambient temperature.
Kung nagawa mong makuha tubig ulan sa sa malaking bilang, diligan ang lahat ng iyong mga halaman gamit lamang ito. Kung hindi, ang mga halaman na sensitibo sa calcium ay dapat na natubigan ng pinalamig na pinakuluang tubig.
Kapag nag-spray ng mga halaman sa tubig mula sa gripo maaaring mangyari na, pagkatapos ng pagpapatayo, ang tubig ay mag-iiwan ng pangit na puting mga spot sa mga dahon; Ito ay mga deposito ng asin. Madali silang matanggal gamit ang malambot na tela o tuyong espongha.
Pagtaas ng kahalumigmigan ng hangin
Maraming mga halaman ang nangangailangan ng mataas na kahalumigmigan, lalo na sa mga tuyo at mahusay na pinainit na mga silid. Samakatuwid, dapat kang lumikha ng isang sapat na mahalumigmig na kapaligiran para sa kanila. Ang isang paraan upang makamit ang layuning ito ay araw-araw na pag-spray ng tubig sa mga dahon mula sa isang maginoo na sprayer. Huwag kalimutan na ang operasyong ito ay dapat gawin sa lilim. Ang isa pang paraan ay ilagay ang mga palayok ng halaman sa isang malawak na tray na puno ng isang layer ng graba o maliliit na bato at natatakpan ng tubig. Ang isa pang paraan upang magbigay ng kahalumigmigan ay ang paglubog ng bawat palayok sa isang mas malaking sisidlan, tulad ng isang pampalamuti na lalagyan. Pagkatapos nito, punan hanggang sa mga gilid ang puwang na nabuo sa pagitan ng dalawang sisidlan na may pit, na dapat na patuloy na panatilihing basa-basa.



